Heathcote Et Al 2012 High Resolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Qadan, the Jebel Sahaba Cemetery and the Lithic
PL ISSN 0066–5924 INDEX 32597x (VAT 8%) DOI: 10.23858/APa58.2020 Institute of Archaeology and Ethnology Polish Academy of Sciences Archaeologia Polona vol. 58: 2020 Special theme: PREHistorY OF North-EAST AFRICA Volume dedicated to Prof. Michał Kobusiewicz on his 80th birthday Archaeologia Polona Volume 58 : 2020 Editor: DAGMARA H. WERRA Associate Editor MARZENA WOŹNY International Advisory Prof. PETER BOGUCKI – Princeton University, USA Board: Prof. MARTIN GOJDA – University of Western Bohemia, Pilsen, Czech Republic Prof. ANTHONY HARDING – University of Exeter, Great Britain Prof. MARIA IACOVOU – University of Cyprus, Nicosia, Cyprus Prof. CAROLA METZNER-NEBELSICK – University of Munich, Germany Prof. BJØRNAR OLSEN – University of Tromsø, Norway Prof. GUIDO VANNINI – University of Florence, Italy Editors of the volume: PRZEMYSłAW Bobrowski and MirosłAW MASOJć Linguistic consultation: PAUL BARFORD Cover illustration: MACIEJ JÓRDECZKA Reviewers: Mirosław Furmanek (Wroclaw), Elena Garcea (Cassino), Maria Gatto (Leicester), Bolesław Ginter (Cracow), Tomasz Herbich (Warsaw), Karla Kroeper (Berlin), Alice Leplongeon (Leuven), Maria Kaczmarek (Poznan), Andrea Manzo (Naples), Arkadiusz Marciniak (Poznan), Henryk Paner (Gdansk), Tomasz Płonka (Wroclaw), Włodzimierz Rączkowski (Poznan), Andrzej Rozwadowski (Poznan), Jiří Svoboda (Brno), Philip Van Peer (Leuven), András Zboray (Budapest). Editorial office: Institute of Archaeology and Ethnology Polish Academy of Sciences, 00-140 Warsaw; Al. Solidarności 105 Tel. (4822) 6202881 ext. 154; Fax. (4822) 624 01 00 E-mail: [email protected] Website: http://journals.iaepan.pl/apolona DOI: 10.23858/APa DOI: 10.23858/APa58.2020 Archaeologia Polona Copyright © 2020 by Institute of Archaeology and Ethnology Polish Academy of Sciences. All articles in this volume are published in an open access under the CC BY 4.0 license (https://creativecommons.org/licenses/by/4.0/). -

A Bioarchaeological Perspective Michael Pietrusewskya, Michele Toomay Douglasa, Marilyn K
This article was downloaded by: [Professor Michael Pietrusewsky] On: 07 November 2014, At: 13:30 Publisher: Routledge Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK The Journal of Island and Coastal Archaeology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/uica20 Health in Ancient Mariana Islanders: A Bioarchaeological Perspective Michael Pietrusewskya, Michele Toomay Douglasa, Marilyn K. Swiftb, Randy A. Harperb & Michael A. Flemingb a Department of Anthropology, University of Hawai‘i at Mānoa, Honolulu, Hawai‘i, USA b Swift and Harper Archaeological Resource Consulting, Saipan, Northern Mariana Islands, USA Published online: 06 Nov 2014. To cite this article: Michael Pietrusewsky, Michele Toomay Douglas, Marilyn K. Swift, Randy A. Harper & Michael A. Fleming (2014) Health in Ancient Mariana Islanders: A Bioarchaeological Perspective, The Journal of Island and Coastal Archaeology, 9:3, 319-340, DOI: 10.1080/15564894.2013.848959 To link to this article: http://dx.doi.org/10.1080/15564894.2013.848959 PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. -

Bus Schedule Carmel Catholic School Agat and Santa Rita Area to Mount Bus No.: B-39 Driver: Salas, Vincent R
BUS SchoolSCHEDULE Year 2020 - 2021 Dispatcher Bus Operations - 646-3122 | Superintendent Franklin F. Tait ano - 646-3208 | Assistant Superintendent Daniel B. Quintanilla - 647-5025 THE DEPARTMENT OF PUBLIC WORKS, BUS OPERATIONS REQUIRES ALL STUDENTS TO WEAR A MASK PRIOR TO BOARDING THE BUS. THERE WILL BE ONE CHILD PER SEAT FOR SOCIAL DISTANCING. PLEASE ANTICIPATE DELAYS IN PICK UP AND DROP OFF AT DESIGNATED BUS SHELTERS. THANK YOU. TENJO VISTA AND SANTA RITA AREAS TO O/C-30 Hanks 5:46 2:29 OCEANVIEW MIDDLE SCHOOL O/C-29 Oceanview Drive 5:44 2:30 A-2 Tenjo Vista Entrance 7:30 4:01 O/C-28 Nimitz Hill Annex 5:40 2:33 A-3 Tenjo Vista Lower 7:31 4:00 SOUTHERN HIGH SCHOOL 6:15 1:50 AGAT A-5 Perez #1 7:35 3:56 PAGACHAO AREA TO MARCIAL SABLAN DRIVER: AGUON, DAVID F. A-14 Lizama Station 7:37 3:54 ELEMENTARY SCHOOL (A.M. ONLY) BUS NO.: B-123 A-15 Borja Station 7:38 3:53 SANTA ANA AREAS TO SOUTHERN HIGH SCHOOL A-38 Pagachao Upper 7:00 A-16 Naval Magazine 7:39 3:52 MARCIAL SABLAN ELEMENTARY SCHOOL 7:10 STATION LOCATION NAME PICK UP DROP OFF A-17 Sgt. Cruz 7:40 3:51 A-44 Tracking Station Entrance 5:50 2:19 A-18 M & R Store 7:41 3:50 PAGACHAO AREA TO OCEANVIEW MIDDLE A-43 Cruz #2 5:52 2:17 SCHOOL A-42 San Nicolas 5:54 2:15 A-19 Annex 7:42 3:49 A-41 Quidachay 5:56 2:12 A-20 Rapolla Station 7:43 3:48 A-46 Round Table 7:15 3:45 A-40 Santa Ana 5:57 2:11 OCEANVIEW MIDDLE SCHOOL 7:50 3:30 A-38 Pagachao Upper 7:22 3:53 A-39 Last Stop 5:59 2:10 A-37 Pagachao Lower 7:25 3:50 SOUTHERN HIGH SCHOOL 6:11 1:50 HARRY S. -

Lukenya Hill, Kenya)
Late Pleistocene age and archaeological context for the hominin calvaria from GvJm-22 (Lukenya Hill, Kenya) Christian A. Tryona,1, Isabelle Crevecoeurb, J. Tyler Faithc, Ravid Ekshtaina, Joelle Nivensd, David Pattersone, Emma N. Mbuaf, and Fred Spoorg,h aDepartment of Anthropology, Harvard University, Cambridge, MA 02138; bUnité Mixte de Recherche 5199, de la Préhistoire à l’Actuel: Culture, Environnement, et Anthropologie, Centre National de la Recherche Scientifique, Université de Bordeaux, 33615 Talence, France; cArchaeology Program, School of Social Science, University of Queensland, Brisbane, QLD 4072, Australia; dDepartment of Anthropology, New York University, New York, NY 10003; eCenter for the Advanced Study of Hominid Paleobiology, Department of Anthropology, The George Washington University, Washington, DC 20052; fNational Museums of Kenya, Nairobi, Kenya 00100; gDepartment of Human Evolution, Max Planck Institute for Evolutionary Anthropology, D-04103, Leipzig, Germany; and hDepartment of Cell and Developmental Biology, University College London, WC1E 6BT London, United Kingdom Edited by Erik Trinkaus, Washington University, St. Louis, MO, and approved January 16, 2015 (received for review September 19, 2014) Kenya National Museums Lukenya Hill Hominid 1 (KNM-LH 1) is Hominid 1 (KNM-LH 1) partial calvaria from site GvJm-22 at a Homo sapiens partial calvaria from site GvJm-22 at Lukenya Hill, Lukenya Hill, Kenya, the only eastern African fossil hominin Kenya, associated with Later Stone Age (LSA) archaeological de- from a Last Glacial Maximum [LGM; 19–26.4 kya (19)] LSA posits. KNM-LH 1 is securely dated to the Late Pleistocene, and archaeological context. We construct a revised accelerator mass samples a time and region important for understanding the origins spectrometry (AMS) radiocarbon chronology built on 26 new of modern human diversity. -

Guam 179: Facing Te New,Pacific Era
, DOCONBOT Busehis BD 103 349 RC 011 911 TITLE Guam 179: Facing te New,Pacific Era. AnnualEconom c. Review. INSTITUTION .Guam Dept. of Commerce, Agana. SPONS AGENCY Department of the Interior, Washington, D.C. PUB DATE Aug 79 . NOTE 167p.: Docugent prepared by the Economic,Research Center. EDRS ?RICE . 1F01/PC07Plus 'Postage. DESCRIPTORS Annual Reports: Business: *Demography: *Economic Dpveloptlent4 Employment Patternp:Expenditures: Federal GoVernment: Financial, Support: *Government Role:_Local Government: *Productivity;_ Vahles (Data); Tourism - IDENTIFIER, *Guam Micronesia A ABSTRACT Socioeconomic conditions and developmentSare analysed.in thiseport, designed to ge. useful\ tb plannersin government and t_vr,. rivate sector. The introduction sunrmarizes Guam's economic olthook emphasizing the eftect of federalfunds for reconstruction folloVing SupertirphAon.Pamela in 1976,moderate growth ,in tour.ism,,and Guam's pqtential to partici:pateas a staging point in trade between the United States and mainlandQhina The body of the report contains populaktion, employment, and incomestatistics; an -account of th(ik economic role of local and federalgovernments and the military:adescription of economic activity in the privatesector (i.e., tour.isid, construction, manufacturingand trarde, agziculture and .fisheries and finan,cial inStitutionsi:and a discussion of onomic development in. Micronesia 'titsa whole.. Appendices contain them' 1979 uGuam Statistical Abstract which "Constitutesthe bulk Of tpe report and provides a wide lia.riety of data relevantto econ9mic development and planning.. Specific topics includedemography, vital statistics, school enrollment, local and federalgovernment finance, public utilities, transportation, tourism, andinternational trade. The most current"data are for fiscalyear 1977 or 197B with many tables showing figures for the previous 10years.(J11) A , . ***************t*********************************************t********* * . -

Freshwater Use Customs on Guam an Exploratory Study
8 2 8 G U 7 9 L.I:-\'I\RY INT.,NATIONAL R[ FOR CO^.: ^,TY W SAMIATJON (IRC) FRESHWATER USE CUSTOMS ON GUAM AN EXPLORATORY STUDY Technical Report No. 8 iei- (;J/O; 8;4J ii ext 141/142 LO: FRESHWATER USE CUSTOMS ON AN EXPLORATORY STUDY Rebecca A. Stephenson, Editor UNIVERSITY OF GUAM Water Resources Research Center Technical Report No. 8 April 1979 Partial Project Completion Report for SOCIOCULTURAL DETERMINANTS OF FRESHWATER USES IN GUAM OWRT Project No. A-009-Guam, Grant Agreement Nos. 14-34-0001-8012,9012 Principal Investigator: Rebecca A- Stephenson Project Period: October 1, 1977 to September 30, 1979 The work upon which this publication is based was supported in part by funds provided by the Office of Water Research and Technology, U. S. Department of the Interior, Washington, D. C, as authorized by the Water Research and Development Act of 1978. T Contents of this publication do not necessarily reflect the views and policies of the Office of Water Research and Technology, U. S. Department of the Interior, nor does mention of trade names or commercial products constitute their endorsement or recommendation for use by the U- S. Government. ii ABSTRACT Traditional Chamorro freshwater use customs on Guam still exist, at least in the recollections of Chamorros above the age of 40, if not in actual practice in the present day. Such customs were analyzed in both their past and present contexts, and are documented to provide possible insights into more effective systems of acquiring and maintain- ing a sufficient supply of freshwater on Guam. -
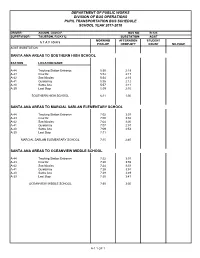
Department of Public Works Division of Bus Operations Pupil Transportation Bus Schedule School Year 2017-2018
DEPARTMENT OF PUBLIC WORKS DIVISION OF BUS OPERATIONS PUPIL TRANSPORTATION BUS SCHEDULE SCHOOL YEAR 2017-2018 DRIVER: AGUON, DAVID F. BUS NO. B-123 SUPERVISOR: TAIJERON, RICKY U. SUBSTATION: AGAT MORNING AFTERNOON STUDENT S T A T I O N S PICK-UP DROP-OFF COUNT MILEAGE AGAT SUBSTATION SANTA ANA AREAS TO SOUTHERN HIGH SCHOOL STATION LOCATION NAME A-44 Tracking Station Entrance 5:50 2:19 A-43 Cruz #2 5:52 2:17 A-42 San Nicolas 5:54 2:15 A-41 Quidachay 5:56 2:12 A-40 Santa Ana 5:57 2:11 A-39 Last Stop 5:59 2:10 SOUTHERN HIGH SCHOOL 6:11 1:50 SANTA ANA AREAS TO MARCIAL SABLAN ELEMENTARY SCHOOL A-44 Tracking Station Entrance 7:02 3:03 A-43 Cruz #2 7:00 3:02 A-42 San Nicolas 7:04 3:00 A-41 Quidachay 7:07 2:57 A-40 Santa Ana 7:09 2:53 A-39 Last Stop 7:11 MARCIAL SABLAN ELEMENTARY SCHOOL 7:15 2:40 SANTA ANA AREAS TO OCEANVIEW MIDDLE SCHOOL A-44 Tracking Station Entrance 7:22 3:57 A-43 Cruz #2 7:20 3:55 A-42 San Nicolas 7:24 3:53 A-41 Quidachay 7:26 3:51 A-40 Santa Ana 7:28 3:49 A-39 Last Stop 7:30 3:47 OCEANVIEW MIDDLE SCHOOL 7:35 3:30 A-1 1 OF 1 DEPARTMENT OF PUBLIC WORKS DIVISION OF BUS OPERATIONS PUPIL TRANSPORTATION BUS SCHEDULE SCHOOL YEAR 2017-2018 DRIVER: BORJA, GARY P. -

UNIVERSITY of HAWAII LIBRARY
UNIVERSITY Of HAWA II LI BRARY 0 RECORDS OF 'IllE GERMAN IMPEHIAL GOVEHNMENT • OF THE SOUTII SEAS PER'D\INING TO MICRONESIA J\S CON'D\ INED IN 'fl-IE A,rtC.:llIVES Ol"F'ICE , AUSTRALIJ\N NATION/\L GOVlmNMENT CANBERHA Volume 8 CRS Gl, ITEM 9-2 General Administration, Caroline Islands 1899 - 1907 Property of l>i.vislon of Lancls ancl Surveys UcparlmcnL of Resources and Dcvclopmcnl Trust Territo ry Government Snipan, Mariana Islands 96950 CRS G1 ITEM 9-~ ALLGE~EINE VER ~ ALTUNG KAROLINEN 1899-1907 ( GEi~3!L.L .rt ~l ..Ii:IL7H.i.T I Oli CA.HOLIHE ISLAirnS) RECORDS OF nrn GERMAN IMPERIAL GOVERNMENT. OF THE SOUTii SFAS PER'D\INING TO MICRONESIA AS CONTAINED IN nrn ARCHIVES OFFICE , AUSTRALIAN NATIONAL GOVERNMENT CANBERRA Volume 8 CRS Gl , ITEM 9-2 General Administration, Caroline Islands 1899 - 1907 Property of Division of Lands and Surveys Department of Resources and Development Trust Territory Government Saipan, Mariana Islands 96950 INTRODUCTION The two main sources for land records and documents relating t~ the Administration of Micronesia by Germany (Marshalls 1885-1914; Carolines and Marianas 1899-1914) are the Commonwealth of Australia Archives Office in Canberra and the Central German Archives at Potsdam.in East Germany • • The German records in Australia were acquired by the Australian Military Administration of New Guinea between 1914 and 1922 from Rabaul, the ' former German capital of German New Guinea and the Islands Sphere (Micronesia). These records are voluminous, and James B. Johnson, Senior Land Commissioner, Mariana Islands District, was sent to Canberra for ten (10) days in August 1969 to examine these records. -

Evolution of Wisdom: Major and Minor Keys
EVOLUTION OF WISDOM: MAJOR AND MINOR KEYS AGUSTÍN FUENTES AND CELIA DEANE-DRUMMOND Center for Theology, Science, and Human Flourishing University of Notre Dame Evolution of Wisdom: Major and Minor Keys by Center for Theology, Science, and Human Flourishing is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, except where otherwise noted. Copyright © 2018 Center for Theology, Science, and Human Flourishing, University of Notre Dame CONTENTS Contents v Acknowledgements vii List of Contributors viii Introduction: Transdisciplinarity, Evolution, and Engaging Wisdom 1 Agustín Fuentes and Celia Deane-Drummond PART I. INTERDISCIPLINARY WISDOM 1. Independent Reason, Faith, and a Distinctively Human Wisdom 7 Angela Carpenter 2. Re-Engaging Theology and Evolutionary Biology: The Nature of True Wisdom 15 Nicola Hoggard Creegan 3. Human Origins and the Emergence of a Distinctively Human Imagination 25 J. Wentzel van Huyssteen PART II. EVOLUTIONARY NARRATIVES 4. Technological Intelligence or Social Wisdom? Promiscuous Sociality, Things, 41 and Networks in Human Evolution Fiona Coward 5. The Palaeolithic Archaeological Record and the Materiality of Imagination: A 57 Response to J. Wentzel van Huyssteen Jennifer French 6. How did Hominins become Human? 64 Marc Kissel PART III. WISDOM AND THE MIND 7. De-Centering Humans within Cognitive Systems 83 Marcus Baynes-Rock 8. Practical Wisdom: Good Reasoning or Good Action? 89 Craig IfGand 9. Concepts of Reason and Wisdom 96 Maureen Junker-Kenny 10. Wisdom and Freedom as Reason - Sensitive Action Control 104 Aku Visala PART IV. WISDOM IN THE MINOR KEY 11. Evolution in the Minor Key 115 Tim Ingold 12. A Response to Tim Ingold: Evolution in the Minor Key 124 Karen Kilby 13. -

Warfare and Social Change in Non-State Societies of the Predynastic Nile Valley
Aula Orientalis. Revista de estudios del Próximo Oriente Antiguo, vol. 33, 2015, pp. 43-49. Warfare and Social Change in non-State Societies of the Predynastic Nile Valley. Augusto Gayubas. Cita: Augusto Gayubas (2015). Warfare and Social Change in non-State Societies of the Predynastic Nile Valley. Aula Orientalis. Revista de estudios del Próximo Oriente Antiguo, 33, 43-49. Dirección estable: https://www.aacademica.org/augusto.gayubas/8 Acta Académica es un proyecto académico sin fines de lucro enmarcado en la iniciativa de acceso abierto. Acta Académica fue creado para facilitar a investigadores de todo el mundo el compartir su producción académica. Para crear un perfil gratuitamente o acceder a otros trabajos visite: https://www.aacademica.org. Warfare and Social Change in Non-state Societies of the Predynastic Nile Valley Augusto Gayubas – Buenos Aires, Argentina CONICET / University of Buenos Aires [The emergence of leadership and chiefdoms in the Predynastic Nile Valley has been a topic of interest among researchers. Authors who have dealt with the issue tend to focus their research mainly on the economic and administrative aspects, lacking a serious consideration of conflict, and particularly warfare. Nevertheless, archaeological testimonies seem to suggest a close relation between the emergence of socio-political hierarchies and the realm of war in the Predynastic Nile Valley. The aim of this paper is to briefly consider this issue by evaluating the available archaeological evidence.] Keywords: warfare, chiefdom, Nile Valley, Predynastic period. 1. The emergence of leadership and chiefdoms in the Nile Valley: consensualist approaches The emergence of leadership and socio-political hierarchy among non-State societies of the Predynastic Nile Valley –that is to say, prior to the emergence of the State– has proved to be a topic of much interest among researchers. -

The Amarna South Tombs Cemetery
The Amarna South Tombs Cemetery: Biocultural Dynamics of a Disembedded Capital City in New Kingdom Egypt by William Charles Schaffer A Dissertation Presented in Partial Fulfillment of the Requirements for the Degree Doctor of Philosophy Approved November 2018 by the Graduate Supervisory Committee: Jane E. Buikstra, Chair Christopher M. Stojanowski Michael E. Smith Jerome C. Rose ARIZONA STATE UNIVERSITY December 2018 ABSTRACT The Egyptian New Kingdom city of Akhetaten (modern: Tell el-Amarna, el- Amarna, or simply Amarna) provides a unique opportunity to study ancient biocultural dynamics. It was a disembedded capital removed from the major power bases of Memphis and Thebes that was built, occupied, and abandoned within approximately 20 years (c. 1352–1336 BCE). This dissertation used the recently excavated Amarna South Tombs cemetery to test competing models for the development of disembedded capitals, such as the geographic origin of its migrants and its demographic structure in comparison to contrastive models for the establishment of settlements. The degree to which biological relatedness organized the South Tombs cemetery was also explored. The results suggest that the Nile Valley into the New Kingdom (1539–1186 BCE) was very diverse in dental cervical phenotype and thus highly mobile in respects to gene flow, failing to reject that the Amarna city was populated by individuals and families throughout the Nile Valley. In comparison, the Amarna South Tombs cemetery contained the least amount of dental phenotypic diversity, supporting a founder effect due to migration from larger, more diverse gene pools to the city or the very fact that the city and sample only reflect a 20- year interval with little time to accumulate phenotypic variation. -

Implementation of the State Children's Health Insurance Program
Contract No.: 500-96-0016(03) MPR Reference No.: 8644-100 Implementation of the State Children’s Health Insurance Program: Synthesis of State Evaluations Background for the Report to Congress March 2003 Margo Rosenbach Marilyn Ellwood Carol Irvin Cheryl Young Wendy Conroy Brian Quinn Megan Kell Submitted to: Submitted by: Centers for Medicare & Medicaid Services Mathematica Policy Research, Inc. Office of Research, Development, 50 Church Street, 4th Floor and Information Cambridge, MA 02138 7500 Security Boulevard (617) 491-7900 Baltimore, MD 21244 Project Officer: Project Director: Rosemarie Hakim Margo Rosenbach This report was prepared for the Centers for Medicare & Medicaid Services, U.S. Department of Health and Human Services, under CMS contract number 500-96-0016 (03). The contents of this publication do not necessarily reflect the views or policies of CMS or DHHS, nor does the mention of trade names, commercial products, or organizations imply endorsement by CMS, DHHS, or Mathematica Policy Research, Inc. The authors are solely responsible for the contents of this publication. CONTENTS Chapter Page EXECUTIVE SUMMARY............................................................................. xiii I INTRODUCTION...............................................................................................1 A. OVERVIEW OF SCHIP AS OF MARCH 31, 2001 ....................................1 B. RATIONALE FOR THIS REPORT .............................................................7 C. ORGANIZATION OF THIS REPORT ........................................................9