Implications for Conservation Helena Reis Batalha
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Global Assessment of Parasite Diversity in Galaxiid Fishes
diversity Article A Global Assessment of Parasite Diversity in Galaxiid Fishes Rachel A. Paterson 1,*, Gustavo P. Viozzi 2, Carlos A. Rauque 2, Verónica R. Flores 2 and Robert Poulin 3 1 The Norwegian Institute for Nature Research, P.O. Box 5685, Torgarden, 7485 Trondheim, Norway 2 Laboratorio de Parasitología, INIBIOMA, CONICET—Universidad Nacional del Comahue, Quintral 1250, San Carlos de Bariloche 8400, Argentina; [email protected] (G.P.V.); [email protected] (C.A.R.); veronicaroxanafl[email protected] (V.R.F.) 3 Department of Zoology, University of Otago, P.O. Box 56, Dunedin 9054, New Zealand; [email protected] * Correspondence: [email protected]; Tel.: +47-481-37-867 Abstract: Free-living species often receive greater conservation attention than the parasites they support, with parasite conservation often being hindered by a lack of parasite biodiversity knowl- edge. This study aimed to determine the current state of knowledge regarding parasites of the Southern Hemisphere freshwater fish family Galaxiidae, in order to identify knowledge gaps to focus future research attention. Specifically, we assessed how galaxiid–parasite knowledge differs among geographic regions in relation to research effort (i.e., number of studies or fish individuals examined, extent of tissue examination, taxonomic resolution), in addition to ecological traits known to influ- ence parasite richness. To date, ~50% of galaxiid species have been examined for parasites, though the majority of studies have focused on single parasite taxa rather than assessing the full diversity of macro- and microparasites. The highest number of parasites were observed from Argentinean galaxiids, and studies in all geographic regions were biased towards the highly abundant and most widely distributed galaxiid species, Galaxias maculatus. -

Dwarf Galaxias
Threatened Species Link www.tas.gov.au SPECIES MANAGEMENT PROFILE Galaxiella pusilla Eastern Dwarf Galaxias Group: Chordata (vertebrates), Actinopterygii (bony fish), Salmoniformes (salmonids), Galaxiidae Status: Threatened Species Protection Act 1995: vulnerable Environment Protection and Biodiversity Conservation Act 1999: Vulnerable Endemic Found in Tasmania and elsewhere Status: A complete species management profile is not currently available for this species. Check for further information on this page and any relevant Activity Advice. Key Points Important: Is this species in your area? Do you need a permit? Ensure you’ve covered all the issues by checking the Planning Ahead page. Important: Different threatened species may have different requirements. For any activity you are considering, read the Activity Advice pages for background information and important advice about managing around the needs of multiple threatened species. Further information Check also for listing statement or notesheet pdf above (below the species image). Recovery Plan Cite as: Threatened Species Section (2021). Galaxiella pusilla (Eastern Dwarf Galaxias): Species Management Profile for Tasmania's Threatened Species Link. https://www.threatenedspecieslink.tas.gov.au/Pages/Dwarf-Galaxias.aspx. Department of Primary Industries, Parks, Water and Environment, Tasmania. Accessed on 29/9/2021. Contact details: Threatened Species Section, Department of Primary Industries, Parks, Water and Environment, GPO Box 44, Hobart, Tasmania, Australia, 7001. Phone (1300 368 550). Permit: A permit is required under the Tasmanian Threatened Species Protection Act 1995 to 'take' (which includes kill, injure, catch, damage, destroy and collect), keep, trade in or process any specimen or products of a listed species. Additional permits may also be required under other Acts or regulations to take, disturb or interfere with any form of wildlife or its products, (e.g. -

AFSS 2014 Abstract Booklet
Oral abstracts Oral abstracts The evolution of ASFB – reflections on 35 years of membership Seeing with sound – the behaviour and movements of fish in estuaries 1 2 1 2 01 Martin Gomon1 04 Alistair Becker , Iain M Suthers , Alan K Whitfield 1. Museum Victoria, Melbourne, VIC, Australia 1. University of New South Wales, Sydney, NSW, Australia 2. South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The Australian Society for Fish Biology had as its inception early meetings organised by the ichthyological staff of the Australian Museum and New South Fisheries intended as a mechanism for sharing advances in fish related science and the development of Underwater video techniques have progressed rapidly over the past ten years, and are now used in a diverse range of habitats initiatives leading to the better understanding of the diversity and biology of Australia’s ichthyofauna. Although the enthusiasm from small creeks to the ocean depths. A limitation of underwater video cameras is they rely on high levels of water clarity and and casual nature of this now incorporated body has remained, the various focuses of the Society and its annual conferences have require artificial lighting if used in low light conditions. In systems such as estuaries, turbidity levels often restrict the use of changed over the decades in line with the evolving directions of the institutions and authorities charged with addressing fish conventional video. Acoustic cameras (DIDSON) overcome this problem as they rely on sound to produce near video, flowing studies and management, as well as the transient influences of the many characters that have been the Society’s driving force. -

A New Subspecies of Eurasian Reed Warbler Acrocephalus Scirpaceus in Egypt
Jens Hering et al. 101 Bull. B.O.C. 2016 136(2) A new subspecies of Eurasian Reed Warbler Acrocephalus scirpaceus in Egypt by Jens Hering, Hans Winkler & Frank D. Steinheimer Received 4 December 2015 Summary.—A new subspecies of European Reed Warbler Acrocephalus scirpaceus is described from the Egypt / Libya border region in the northern Sahara. Intensive studies revealed the new form to be clearly diagnosable within the Eurasian / African Reed Warbler superspecies, especially in biometrics, habitat, breeding biology and behaviour. The range of this sedentary form lies entirely below sea level, in the large depressions of the eastern Libyan Desert, in Qattara, Siwa, Sitra and Al Jaghbub. The most important field characters are the short wings and tarsi, which are significantly different from closely related A. s. scirpaceus, A. s. fuscus and A. s. avicenniae, less so from A. baeticatus cinnamomeus, which is more clearly separated by behaviour / nest sites and toe length. Molecular genetic analyses determined that uncorrected distances to A. s. scirpaceus are 1.0–1.3%, to avicenniae 1.1–1.5% and to fuscus 0.3–1.2%. The song is similar to that of other Eurasian Reed Warbler taxa as well as that of African Reed Warbler A. baeticatus, but the succession of individual elements appears slower than in A. s. scirpaceus and therefore shows more resemblance to A. s. avicenniae. Among the new subspecies’ unique traits are that its preferred breeding habitat in the Siwa Oasis complex, besides stands of reed, is date palms and olive trees. A breeding density of 107 territories per 10 ha was recorded in the cultivated area. -

AERC Wplist July 2015
AERC Western Palearctic list, July 2015 About the list: 1) The limits of the Western Palearctic region follow for convenience the limits defined in the “Birds of the Western Palearctic” (BWP) series (Oxford University Press). 2) The AERC WP list follows the systematics of Voous (1973; 1977a; 1977b) modified by the changes listed in the AERC TAC systematic recommendations published online on the AERC web site. For species not in Voous (a few introduced or accidental species) the default systematics is the IOC world bird list. 3) Only species either admitted into an "official" national list (for countries with a national avifaunistic commission or national rarities committee) or whose occurrence in the WP has been published in detail (description or photo and circumstances allowing review of the evidence, usually in a journal) have been admitted on the list. Category D species have not been admitted. 4) The information in the "remarks" column is by no mean exhaustive. It is aimed at providing some supporting information for the species whose status on the WP list is less well known than average. This is obviously a subjective criterion. Citation: Crochet P.-A., Joynt G. (2015). AERC list of Western Palearctic birds. July 2015 version. Available at http://www.aerc.eu/tac.html Families Voous sequence 2015 INTERNATIONAL ENGLISH NAME SCIENTIFIC NAME remarks changes since last edition ORDER STRUTHIONIFORMES OSTRICHES Family Struthionidae Ostrich Struthio camelus ORDER ANSERIFORMES DUCKS, GEESE, SWANS Family Anatidae Fulvous Whistling Duck Dendrocygna bicolor cat. A/D in Morocco (flock of 11-12 suggesting natural vagrancy, hence accepted here) Lesser Whistling Duck Dendrocygna javanica cat. -

Deficiencies in Our Understanding of the Hydro-Ecology of Several Native Australian Fish: a Rapid Evidence Synthesis
Marine and Freshwater Research, 2018, 69, 1208–1221 © CSIRO 2018 https://doi.org/10.1071/MF17241 Supplementary material Deficiencies in our understanding of the hydro-ecology of several native Australian fish: a rapid evidence synthesis Kimberly A. MillerA,D, Roser Casas-MuletB,A, Siobhan C. de LittleA, Michael J. StewardsonA, Wayne M. KosterC and J. Angus WebbA,E ADepartment of Infrastructure Engineering, The University of Melbourne, Parkville, Vic. 3010, Australia. BWater Research Institute, Cardiff University, The Sir Martin Evans Building, Museum Avenue, Cardiff, CF10 3AX, UK. CArthur Rylah Institute for Environmental Research, Department of Environment, Land, Water and Planning, Heidelberg, Vic. 3084, Australia. DPresent address: Healesville Sanctuary, Badger Creek Road, Healesville, Vic. 3777, Australia. ECorresponding author. Email address: [email protected] Page 1 of 30 Marine and Freshwater Research © CSIRO 2018 https://doi.org/10.1071/MF17241 Table S1. All papers located by standardised searches and following citation trails for the two rapid evidence assessments All papers are marked as Relevant or Irrelevant based on a reading of the title and abstract. Those deemed relevant on the first screen are marked as Relevant or Irrelevant based on a full assessment of the reference.The table contains incomplete citation details for a number of irrelevant papers. The information provided is as returned from the different evidence databases. Given that these references were not relevant to our review, we have not sought out the full citation details. Source Reference Relevance Relevance (based on title (after reading and abstract) full text) Pygmy perch & carp gudgeons Search hit Anon (1998) Soy protein-based formulas: recommendations for use in infant feeding. -

Aspects of the Phylogeny, Biogeography and Taxonomy of Galaxioid Fishes
Aspects of the phylogeny, biogeography and taxonomy of galaxioid fishes Jonathan Michael Waters, BSc. (Hons.) Submitted in fulfilment of the requirements for the degree of Doctor of Philosophy, / 2- Oo ( 01 f University of Tasmania (August, 1996) Paragalaxias dissim1/is (Regan); illustrated by David Crook Statements I declare that this thesis contains no material which has been accepted for the award of any other degree or diploma in any tertiary institution and, to the best of my knowledge and belief, this thesis contains no material previously published o:r written by another person, except where due reference is made in the text. This thesis is not to be made available for loan or copying for two years following the date this statement is signed. Following that time the thesis may be made available for loan and limited copying in accordance with the Copyright Act 1968. Signed Summary This study used two distinct methods to infer phylogenetic relationships of members of the Galaxioidea. The first approach involved direct sequencing of mitochondrial DNA to produce a molecular phylogeny. Secondly, a thorough osteological study of the galaxiines was the basis of a cladistic analysis to produce a morphological phylogeny. Phylogenetic analysis of 303 base pairs of mitochondrial cytochrome b _supported the monophyly of Neochanna, Paragalaxias and Galaxiella. This gene also reinforced recognised groups such as Galaxias truttaceus-G. auratus and G. fasciatus-G. argenteus. In a previously unrecognised grouping, Galaxias olidus and G. parvus were united as a sister clade to Paragalaxias. In addition, Nesogalaxias neocaledonicus and G. paucispondylus were included in a clade containing G. -
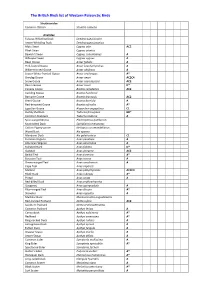
BB List 2016V1
The British Birds list of Western Palearctic Birds Struthionidae Common Ostrich Struthio camelus Anatidae Fulvous Whistling Duck Dendrocygna bicolor Lesser Whistling Duck Dendrocygna javanica Mute Swan Cygnus olor AC2 Black Swan Cygnus atratus Bewick’s Swan Cygnus columbianus A Whooper Swan Cygnus cygnus A Bean Goose Anser fabalis A Pink-footed Goose Anser brachyrhynchus A White-fronted Goose Anser albifrons A Lesser White-fronted Goose Anser erythropus A* Greylag Goose Anser anser AC2C4 Snow Goose Anser caerulescens AC2 Ross’s Goose Anser rossii D* Canada Goose Branta canadensis AC2 Cackling Goose Branta hutchinsii Barnacle Goose Branta leucopsis AC2 Brent Goose Branta bernicla A Red-breasted Goose Branta ruficollis A* Egyptian Goose Alopochen aegyptiaca C1 Ruddy Shelduck Tadorna ferruginea BD* Common Shelduck Tadorna tadorna A Spur-winged Goose Plectropterus gambensis Knob-billed Duck Sarkidiornis melanotos Cotton Pygmy-goose Nettapus coromandelianus Wood Duck Aix sponsa Mandarin Duck Aix galericulata C1 Eurasian Wigeon Anas penelope A American Wigeon Anas americana A Falcated Duck Anas falcata D* Gadwall Anas strepera AC2 Baikal Teal Anas formosa A* Eurasian Teal Anas crecca A Green-winged Teal Anas carolinensis A Cape Teal Anas capensis Mallard Anas platyrhynchos AC2C4 Black Duck Anas rubripes A* Pintail Anas acuta A Red-billed Duck Anas erythrorhyncha Garganey Anas querquedula A Blue-winged Teal Anas discors A* Shoveler Anas clypeata A Marbled Duck Marmaronetta angustirostris Red-crested Pochard Netta rufina AC2 Southern Pochard -

Cape Verde Warbler Research
CONSERVATION, ECOLOGY AND GENETICS OF THE CAPE VERDE WARBLER Acrocephalus brevipennis Report for African Bird Club on fieldwork in Cape Verde November 2013 to January 2014 © Josh Jenkins Shaw Helena Reis Batalha University of East Anglia April 2014 Project details Supervisors Dr. David S. Richardson (Primary supervisor, UEA); Dr. Iain Barr (Co-supervisor, UEA); Dr. Nigel J. Collar (Case partner, BirdLife International); Dr. Paul F. Donald (Advisor, RSPB). Team leader and report writer Helena Reis Batalha Centre for Ecology, Evolution and Conservation School of Biological Sciences University of East Anglia Norwich Research Park NR4 7TJ Norwich United Kingdom Field assistants Jaelsa Mira Moreira (Cape Verde), Andrew Power (Ireland), Josh Jenkins Shaw (United Kingdom) & Jeroen Arnoys (Belgium). Citation Batalha, H. (2014). Conservation, ecology and genetics of the Cape Verde warbler Acrocephalus brevipennis: Report for African Bird Club on fieldwork in Cape Verde, November 2013 to January 2014. Unpublished report, University of East Anglia, Norwich, UK. Work done under the permit 36/2013 issued by the General Direction for the Environment, Achada Santo António, CP 332-A, Praia, Cape Verde Republic. April 2014 Funding and support Table of Contents Project details ........................................................................................................................... 2 Funding and support ............................................................................................................... 2 1. Introduction and -

Cabo Verde Ramsar Information Sheet Published on 18 November 2016 Update Version, Previously Published on 18 July 2005
RIS for Site no. 1577, Lagoa de Pedra Badejo, Cabo Verde Ramsar Information Sheet Published on 18 November 2016 Update version, previously published on 18 July 2005 Cabo Verde Lagoa de Pedra Badejo Designation date 18 July 2005 Site number 1577 Coordinates 15°05'59"N 23°32'12"W Area 666,07 ha https://rsis.ramsar.org/ris/1577 Created by RSIS V.1.7 on - 1 December 2016 RIS for Site no. 1577, Lagoa de Pedra Badejo, Cabo Verde Color codes Fields back-shaded in light blue relate to data and information required only for RIS updates. Note that some fields concerning aspects of Part 3, the Ecological Character Description of the RIS (tinted in purple), are not expected to be completed as part of a standard RIS, but are included for completeness so as to provide the requested consistency between the RIS and the format of a ‘full’ Ecological Character Description, as adopted in Resolution X.15 (2008). If a Contracting Party does have information available that is relevant to these fields (for example from a national format Ecological Character Description) it may, if it wishes to, include information in these additional fields. 1 - Summary Summary The Lagoas de Pedro Badejo wetland is made up of two coastal lagoons on the estuary of two watercourses and the entire basin of one of the watercourses as far as the Poilão reservoir. The lagoons represent an environment of high ecological value for birds, since they contain fresh or slightly brackish water. Since it was flooded, this reservoir has also become an exceptional area for migratory waterbirds and birds endemic to Cape Verde such as the Cape Verde heron (Ardea bournei ) and the Cape Verde swamp-warbler (Acrocephalus brevipennis). -

Acrocephalus and Hippolais Warblers from the Western Palearctic David T
Species limits in Acrocephalus and Hippolais warblers from the Western Palearctic David T. Parkin, Martin Collinson, Andreas J. Helbig, Alan G. Knox, George Sangster and Lars Svensson Sedge Warbler Acrocephalus schoenobaenus Richard Johnson ABSTRACT The taxonomic affiliations within the genera Acrocephalus and Hippolais have long been a matter for debate. Recent molecular and behavioural studies have provided a wealth of new data which can be used to analyse the evolutionary relationships of the Palearctic taxa in these genera. In this paper, we make a series of recommendations for changes in species limits, highlight some problem areas and discuss situations where more research is needed. 276 © British Birds 97 • June 2004 • 276-299 Species limits in Acrocephalus and Hippolais warblers Introduction dering array of small and difficult birds (for long period of taxonomic stability fol- example Emei Leaf Warbler Phylloscopus lowed the publication of the ‘Voous List’ emeiensis, Alström & Olsson 1995; the chiffchaff Afor Holarctic birds (Voous 1977), but in complex, Helbig et al. 1996; and the Greenish recent years there have been dramatic advances Warbler P. trochiloides, Irwin et al. 2001a,b,c), in our knowledge of the evolutionary relation- which has led to an improved understanding of ships of birds, leading to a period of activity their evolution, and hence their taxonomic rela- that shows no sign of abating. These advances tionships. This paper draws together some stem largely from new and exciting ways to recent studies which together provide a clearer study birds, both in the field and in the labora- picture of an especially complex group, the tory. -

A Guide to the Management of Native Fish: Victorian Coastal Rivers and Wetlands 2007
A guide to the management of native fish: Victorian Coastal Rivers and Wetlands 2007 A Guide to the Management of Native Fish: Victorian Coastal Rivers, Estuaries and Wetlands ACKNOWLEDGEMENTS This guide was prepared with the guidance and support of a Steering Committee, Scientific Advisory Group and an Independent Advisory Panel. Steering Committee – Nick McCristal (Chair- Corangamite CMA), Melody Jane (Glenelg Hopkins CMA), Kylie Bishop (Glenelg Hopkins CMA), Greg Peters (Corangamite CMA and subsequently Independent Consultant), Hannah Pexton (Melbourne Water), Rhys Coleman (Melbourne Water), Mark Smith (Port Phillip and Westernport CMA), Kylie Debono (West Gippsland CMA), Michelle Dickson (West Gippsland CMA), Sean Phillipson (East Gippsland CMA), Rex Candy (East Gippsland CMA), Pam Robinson (Australian Government NRM, Victorian Team), Karen Weaver (DPI Fisheries and subsequently DSE, Biodiversity and Ecosystem Services), Dr Jeremy Hindell (DPI Fisheries and subsequently DSE ARI), Dr Murray MacDonald (DPI Fisheries), Ben Bowman (DPI Fisheries) Paul Bennett (DSE Water Sector), Paulo Lay (DSE Water Sector) Bill O’Connor (DSE Biodiversity & Ecosystem Services), Sarina Loo (DSE Water Sector). Scientific Advisory Group – Dr John Koehn (DSE, ARI), Tarmo Raadik (DSE ARI), Dr Jeremy Hindell (DPI Fisheries and subsequently DSE ARI), Tom Ryan (Independent Consultant), and Stephen Saddlier (DSE ARI). Independent Advisory Panel – Jim Barrett (Murray-Darling Basin Commission Native Fish Strategy), Dr Terry Hillman (Independent Consultant), and Adrian Wells (Murray-Darling Basin Commission Native Fish Strategy-Community Stakeholder Taskforce). Guidance was also provided in a number of regional workshops attended by Native Fish Australia, VRFish, DSE, CMAs, Parks Victoria, EPA, Fishcare, Yarra River Keepers, DPI Fisheries, coastal boards, regional water authorities and councils.