Opponens Pollicis Mechanical Effectiveness in Neandertals and Early Modern Humans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

And Thoracic Outlet Syndrome
• Palpatory diagnosis and manipulative management of carpal tunnel syndrome: Part 2. 'Double crush' and thoracic outlet syndrome BENJAMIN M. SUCHER, DO 1( The physician treating carpal nificant. Ultimately, palpatory assessment was tunnel syndrome needs to be aware of the instrumental in guiding the author with initial or possible concomitant occurrence of thoracic subsequent methods (or both) of effective treat outlet syndrome, the so-called double crush syn ment. Palpatory monitoring was the key to clinical drome. Palpation is used to differentiate carpal management in all cases. tunnel syndrome from thoracic outlet syn drome. Such palpatory examination assists Methods the physician in planning the initial treat Patients with CTS were assessed as previously described.? ment, including osteopathic manipulation They all underwent electrodiagnostic testing, which and self-stretching maneuvers, targeted specif included a minimum of median and ulnar distal motor ically at the most clinically significant patho and sensory conduction studies. Needle electromyograms logic region. Supplemental physical medicine and more extensive conduction studies were also per formed if not done previously, or as clinically indicated. modalities such as ultrasound may enhance They were treated according to the outlined protocols the treatment response. Some illustrative for osteopathic manipulation and self-stretching exer cases are reported. cises.B,9 Palpatory assessment routinely included axial rota (Key words: Carpal tunnel syndrome, osteo tion. When restriction was noted for this motion, treat pathic manipulation, thoracic outlet syn ment included the "opponens roll"? technique. As a drome, double crush syndrome) self-stretch maneuver, the thumb is abducted with slight extension and rotated laterally (Figure 1). The primary The initial presentation of carpal tunnel syn limitations or precautions to this new self-stretch involve advanced degenerative changes in the first carpometacarpal drome (CTS) often is a diagnostic challenge, espe joint and bilateral CTS. -

Structural Kinesiology Class 19 Clearing Scars
STRUCTURAL KINESIOLOGY CLASS 19 With John Maguire WHAT WE WILL COVER IN THIS CLASS How to test and clear scars using three approaches: Upper Limb Muscles • Stroking • Teres Major • Oil & Stretch • Coracobrachialis • Figure 8’s • Brachiordialis • Triceps • Opponens Policis 2 CLEARING SCARS About 75% of scars cause an indicator change when Circuit Located (CL’ed). This means there is some energy block through the scar tissue and it has lost its ability to properly transmit chi. Testing to find out if a scar needs clearing: 1. While either you or the person being tested touches the scar, test a previously strong IM. If the scar CL’s, showing indicator weakening, one of the three procedures on the following pages may help. 2. If there is no IM change, touch the frontal eminences and recheck the scar to see if it is hidden. 3. Injury Recall Technique (IRT) may be needed if an accident was involved, which was covered in Class 1. 3 SCAR CORRECTIONS To determine which corrections to use, CL the scar and state: 1. “Stroking” (test) 2. “Oil and Stretch” (test) 3. “Figure 8’s” (test) The one which strengthens the indicator muscle (IM) is the one to use. 4 STROKING SCAR CORRECTION Stroking: 1. Lightly stroke across the scar to find the weakening direction. 2. Find the phase of breath which strengthens (usually the inhalation). 3. Stroke several times in the weaken direction with the strong breath. Work the full length of the scar. 4. Ask if there is a strong emotion related to the scar and if so, have the person hold their frontal eminences and think about the emotion while the correction is being made. -

Deep Dry Needling of the Arm and Hand Muscles 8
Deep dry needling of the arm and hand muscles 8 César Fernández-de-las-Peñas Javier González Iglesias Christian Gröbli Ricky Weissmann CHAPTER CONTENT conditions. Symptoms in the upper quadrant, including the neck, shoulder, arm, forearm, or Introduction . 107 hand not related to an acute trauma or underly- Clinical relevance of TrPs in arm ing systemic diseases, can be provoked by trigger and hand pain syndromes . 108 points (TrPs). In fact, there are several neck and Dry needling of the arm and hand muscles . 108 shoulder muscles with referred pain pattern being perceived throughout the upper extremity, e.g. Coracobrachialis muscle. 108 the scalenes, subclavius, pectoralis minor, supra- Biceps brachii muscle (short head) . 109 spinatus, infraspinatus, subscapularis, pectoralis Triceps brachii muscle (lower portion) . 109 major, latissimus dorsi, serratus posterior supe- Anconeus muscle . 110 rior and serratus anterior muscles ( Simons et al. Brachialis muscle . 110 1999 ). For instance, Qerama et al. (2009) dem- Brachioradialis muscle . 111 onstrated that 49% of individuals with normal electrophysiological findings in the median nerve, Supinator muscle . 111 but with symptoms mimicking carpal tunnel syn- Wrist and fi nger extensor muscles. 112 drome, presented with active TrPs in the infra- Pronator teres muscle . 113 spinatus muscle with paresthesia and referred Wrist and fi nger fl exor muscles . 113 pain to the arm and fingers. In the same study, Flexor pollicis longus, extensor pollicis patients with mild electrophysiological signs of longus, and abductor pollicis longus . 114 carpal tunnel syndrome exhibited a significantly Extensor indicis muscle . 115 higher occurrence of infraspinatus muscle TrPs in the symptomatic arm as compared with patients Adductor pollicis, opponens pollicis, with moderate to severe electrophysiological fl exor pollicis brevis, and abductor pollicis brevis muscles . -
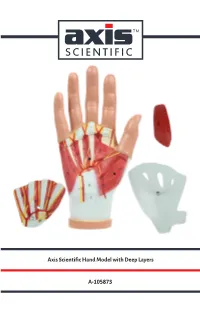
Axis Scientific Hand Model with Deep Layers A-105873
Axis Scientific Hand Model with Deep Layers A-105873 17. Abductor Digiti Minimi Muscle 03. Abductor Pollicis Brevis 01. Palmar Aponeurosis Muscle 03. Abductor Pollicis Brevis Muscle 06. Adductor 18. Flexor Digiti Minimi Brevis Muscle Pollicis Muscle 02. Palmaris Brevis Muscle 11. Ulnar Nerve 21. Palmar Interosseous 12. Ulnar Artery Muscles 22. Palmar 07. Tendon of Palmaris Metacarpal Longus Arteries 20. Deep Palmar (Arterial) Arch 19. Opponens Digiti 04. Flexor Pollicis Brevis Muscle Minimi Muscle 05. Opponens Pollicis Muscle 18. Flexor Digiti Minimi Brevis Muscle 10. Lumbrical 28. Dorsal Interosseous Muscles Muscles 16. Superficial 26. Dorsal Venous Network Palmar Branch of 23. Superficial Branch of Palmar Nerve Radial Artery 25. Radial Artery 24. Dorsal Branch of Ulnar Nerve 27. Extensor Digitorum Tendon 14. 08. Transverse 13. Superficial Median 15. Common Carpal Palmar Arch Nerve Palmar Digital Ligament Nerves and Arteries 07. Tendon 09. Flexor Digitorum of Palmaris Superficialis Tendons Longus 15. Common Palmar Digital 01. Palmar Aponeurosis Nerves and Arteries 16. Superficial Palmar 02. Palmaris Brevis Muscle Branch of Radial Artery 03. Abductor Pollicis Brevis Muscle 17. Abductor Digiti Minimi Muscle 18. Flexor Digiti 04. Flexor Pollicis Brevis Muscle Minimi Brevis Muscle 19. Opponens Digiti 05. Opponens Pollicis Muscle Minimi Muscle 06. Adductor Pollicis Muscle 20. Deep Palmar (Arterial) Arch 07. Tendon of Palmaris Longus 21. Palmar Interosseous Muscles 08. Transverse Carpal Ligament 22. Palmar Metacarpal Arteries 09. Flexor Digitorum Superficialis 23. Superficial Branch of Tendons Palmar Nerve 10. Lumbrical Muscle 24. Dorsal Branch of Ulnar Nerve 11. Ulnar Nerve 25. Radial Artery 26. Dorsal Venous 12. Ulnar Artery Network of Hand 13. -

Anatomical Variations of the Abductor Pollicis Longus: a Pilot Study P
Folia Morphol. Vol. 79, No. 4, pp. 817–822 DOI: 10.5603/FM.a2019.0134 O R I G I N A L A R T I C L E Copyright © 2020 Via Medica ISSN 0015–5659 journals.viamedica.pl Anatomical variations of the abductor pollicis longus: a pilot study P. Karauda1, Ł. Olewnik1, M. Podgórski2, M. Polguj1, K. Ruzik1, B. Szewczyk3, M. Topol1 1Department of Normal and Clinical Anatomy, Interfaculty Chair of Anatomy and Histology, Medical University of Lodz, Poland 2Department of Diagnostic Imaging, Polish Mother’s Memorial Hospital Research Institute, Lodz, Poland 3Department of Clinical Morphology, Medical University of Lodz, Poland [Received: 12 October 2019; Accepted: 3 November 2019] Background: The abductor pollicis longus (APL) originates from the lateral part of the dorsal surface of the body of the ulna below the insertion of the anconeus muscle, from the interosseous membrane, and from the middle third of the dorsal surface of the body of the radius. However, the number of its accessory bands and their insertion vary considerably. Materials and methods: Fifty upper limbs (2 paired, 31 male, 19 female) were obtained from adult Caucasian cadavers, and fixed in 10% formalin solution before examination. Results: The APL muscle was present in all specimens. The muscles were divided into three main categories, with type II and III being dived into subtypes. Type I was characterised by a single distal attachment, with the tendon inserting to the base of the I metacarpal bone. Type II was characterised by a bifurcated distal attachment, with the main tendon inserting to the base of the first metacarpal bone; this type was divided into three subtypes (a–c). -

Supplementary Material for Geometrical Parameters for Musculoskeletal Modeling of Hand
Supplementary material for Geometrical parameters for musculoskeletal modeling of hand L. Havelkov´a,T. Z´ıtka, P. Fiala, M. Rybarova, R. Tup´y,V. Kalis, Ismail K. M. January 30, 2021 This material supplements article Hand muscles attachments: A Geometrical model and serves as dataset documentation as well. It summarizes data collected and describes in detail methods used. The dataset consists of two separate datasets: morphological and morphometric dataset and dataset of positions of muscle lines origins, insertions and CSAs points. Methods described include Matlab code used to calculate lines end- and via-points and details on extractions of origin, insertion and CSA surfaces. Contents 1 Morphological and morphometric dataset based on dissections1 2 Placing muscle action lines8 3 Dataset of positions of muscle lines origins, insertions and CSAs 10 4 Matlab code for calculating lines end and via points 14 1 Morphological and morphometric dataset based on dis- sections The morphological and morphometric dataset consist of morphological drawings (Figure1 to4) and tables1 to4 of morphometric dimensions available in xlsx files. In Figure1 trajectories and attachments of short flexors of thenar and hypothenar groups are shown, the abductor pollicis brevis is in the most lateral position of the thenar group and covers the opponens pollicis muscle. The adductor pollicis is the deepest and the most medial muscle of this group. The two heads (superficial and deep) of the flexor pollicis brevis are located at the centre of the group, separated by the tendon of flexor pollicis longus descending from the forearm. In Figure2 the origins, insertions and CSAs of the opponens pollicis and opponens digiti minimi are depicted. -

Hypertrophied Reverse Palmaris Longus Muscle: a Rare Cause of Carpal Tunnel Syndrome
International Surgery Journal Sundaramurthy N et al. Int Surg J. 2020 Feb;7(2):576-580 http://www.ijsurgery.com pISSN 2349-3305 | eISSN 2349-2902 DOI: http://dx.doi.org/10.18203/2349-2902.isj20200319 Case Report Hypertrophied reverse palmaris longus muscle: a rare cause of carpal tunnel syndrome Narayanamurthy Sundaramurthy, Surya Rao Rao Venkata Mahipathy*, Alagar Raja Durairaj Department of Plastic and Reconstructive Surgery, Saveetha Medical College and Hospital, Thandalam, Kanchipuram, Tamil Nadu, India Received: 04 December 2019 Accepted: 08 January 2020 *Correspondence: Dr. Surya Rao Rao Venkata Mahipathy, E-mail: [email protected] Copyright: © the author(s), publisher and licensee Medip Academy. This is an open-access article distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. ABSTRACT Carpal tunnel syndrome (CTS) is usually secondary to compression or irritation of the median nerve in the fibro- osseous canal formed by the flexor retinaculum (transverse carpal ligament) and the carpal bones. The prevalence of CTS in the general population is about 7 to 19%. Several causes both local and systemic have been described, but CTS due to aberrant musculature are rare. Here we report a case of a middle-aged female with paresthesia of the hand and a positive Phalen’s test with nerve conduction study of the median nerve showing sensorimotor neuropathy. The patient underwent surgery for open CTS release where we found a hypertrophied reverse palmaris longus muscle attached to the palmar aponeurosis which was excised along with its proximal tendon. -

Evidence of a False Thumb in a Fossil Carnivore Clarifies the Evolution of Pandas
Evidence of a false thumb in a fossil carnivore clarifies the evolution of pandas Manuel J. Salesa*†, Mauricio Anto´ n*, Ste´ phane Peigne´ ‡, and Jorge Morales* *Departamento de Paleobiologı´a,Museo Nacional de Ciencias Naturales, Consejo Superior de Investigaciones Cientificas, Jose´Gutie´rrez Abascal 2, 28006 Madrid, Spain; and ‡Laboratoire de Ge´obiologie, Biochronologie et Pale´ontologie Humaine, Centre National de la Recherche Scientifique Unite´Mixte de Recherche 6046, Universite´de Poitiers, 40 Avenue du Recteur Pineau, 86022 Poitiers, France Edited by F. Clark Howell, University of California, Berkeley, CA, and approved October 7, 2005 (received for review June 12, 2005) The ‘‘false thumb’’ of pandas is a carpal bone, the radial sesamoid, and thus uncommon in larger, terrestrial carnivores. Simocyon, which has been enlarged and functions as an opposable thumb. If about the size of a puma, is the largest known arctoid carnivore the giant panda (Ailuropoda melanoleuca) and the red panda with such lumbar morphology, likely reflecting a scansorial (Ailurus fulgens) are not closely related, their sharing of this habit, meaning that the animal would forage on the ground but adaptation implies a remarkable convergence. The discovery of would readily climb when necessary (5). Many features of the previously unknown postcranial remains of a Miocene red panda appendicular skeleton indicate climbing abilities and the lack of relative, Simocyon batalleri, from the Spanish site of Batallones-1 cursorial adaptations. The scapula has a large process for the (Madrid), now shows that this animal had a false thumb. The radial teres major muscle, shared only with the arboreal kinkajou sesamoid of S. -

Patterns of Reinnervation and Motor Unit Recruitment in Human Hand Muscles After Complete Ulnar and Median Nerve Section and Resuture
Journal of Neurology, Neurosurgery, and Psychiatry 1987;50:259-268 Patterns of reinnervation and motor unit recruitment in human hand muscles after complete ulnar and median nerve section and resuture CHRISTINE K THOMAS, RICHARD B STEIN, TESSA GORDON,* ROBERT G LEE,t M GEORGE ELLEKERt From the Departments of Physiology, Pharmacology* and Medicine,1 University of Alberta, Edmonton, and Department ofClinical Neurosciences,j University of Calgary, Calgary, Canada SUMMARY Following complete ulnar or above-elbow median nerve sections, there was no significant correlation between motor unit size (twitch amplitude) and recruitment threshold, as assessed by spike triggered averaging. This absence of orderly recruitment was attributed to misdi- rection of motor axons during regeneration. Following median nerve section at wrist level, where the reinnervated muscles have more synergistic actions, orderly recruitment by size appeared to be re-established. Thus, the size principle of motor unit recruitment can be re-established after nerve section in humans, if motor axons innervate their original muscles or ones with closely synergistic functions. If we want to tie a shoelace, write a letter, eat, or play in the periphery.6 These axons had opportunities to a musical instrument, we are confronted with the need reinnervate their original or foreign muscles. There- to control our hand muscles precisely. Our ability to fore, the disordered recruitment of reinnervated co-ordinate these and many other fine movements is motor units could have resulted from misdirection of generally assumed to result from direct corticospinal motor axons to foreign muscles with different func- connections to the motoneurons innervating the hand tions and/or from changes in the activity of the muscles' 2 and from an orderly recruitment of motoneurons. -

Vesalius on the Hand and Forearm Muscles
UvA-DARE (Digital Academic Repository) On the innovative genius of Andreas Vesalius Brinkman, R.J.C. Publication date 2017 Document Version Other version License Other Link to publication Citation for published version (APA): Brinkman, R. J. C. (2017). On the innovative genius of Andreas Vesalius. General rights It is not permitted to download or to forward/distribute the text or part of it without the consent of the author(s) and/or copyright holder(s), other than for strictly personal, individual use, unless the work is under an open content license (like Creative Commons). Disclaimer/Complaints regulations If you believe that digital publication of certain material infringes any of your rights or (privacy) interests, please let the Library know, stating your reasons. In case of a legitimate complaint, the Library will make the material inaccessible and/or remove it from the website. Please Ask the Library: https://uba.uva.nl/en/contact, or a letter to: Library of the University of Amsterdam, Secretariat, Singel 425, 1012 WP Amsterdam, The Netherlands. You will be contacted as soon as possible. UvA-DARE is a service provided by the library of the University of Amsterdam (https://dare.uva.nl) Download date:06 Oct 2021 Chapter 2 Vesalius on the hand and forearm muscles Chapter based on article: Andreas Vesalius’ 500th anniversary: The initiation of hand and forearm myology Brinkman R.J., Hage J.J. J Hand Surg Eur Vol. 2015;40(9):987-94 17 Chapter 2. Vesalius on the hand and forearm muscles Introduction “When doctors supposed that only the care of internal complaints concerned them, consider- ing a mere knowledge of the viscera as more than enough for them, they neglected the structure of the bones and muscles, as well as of the nerves, veins and arteries that run through bones and muscles, as of no importance for them” [1]. -

Multiple Variations of the Tendons of the Anatomical Snuffbox
Singapore Med J 2014; 55(1): 37-40 O riginal A rticle doi:10.11622/smedj.2013216 Multiple variations of the tendons of the anatomical snuffbox San San Thwin1, MBBS, MMedSc, Fazlin Zaini1, MMed Sc, Myo Than1, MBBS, MMedSc Introduction Multiple tendons of the abductor pollicis longus (APL) in the anatomical snuffbox of the wrist can lead to the development of de Quervain’s syndrome, which is caused by stenosing tenosynovitis. A cadaveric study was performed to establish the variations present in the tendons of the anatomical snuffbox in a Malaysian population, in the hope that this knowledge would aid clinical investigation and surgical treatment of de Quervain’s tenosynovitis. Methods Routine dissection of ten upper limbs was performed to determine the variations in the tendons of the anatomical snuffbox of the wrist. Results In all the dissected upper limbs, the APL tendon of the first extensor compartment was found to have several (3–14) tendon slips. The insertion of the APL tendon slips in all upper limbs were at the base of the first metacarpal bone, trapezium and fascia of the opponens pollicis muscle; however, in seven specimens, they were also found to be attached to the fleshy belly of the abductor pollicis brevis muscle. In two specimens, double tendons of the extensor pollicis longus located in the third extensor compartment were inserted into the capsule of the proximal interphalangeal joints before being joined to the extensor expansion. In two other specimens, the first extensor compartment had two osseofibrous tunnels divided by a septum that separated the APL tendon from the extensor pollicis brevis tendon. -

Axis Scientific 27-Part Half Life-Size Muscular Figure A-105165
Axis Scientific 27-Part Half Life-Size Muscular Figure A-105165 I. Region of Head and Neck 17. Masseter Muscle 1.Frontal Belly of Occipitofrontalis 18. Temporalis Muscle Muscle 19. Lateral Pterygoid Muscle 2. Epieranial Aponeurosis 20. Medial Pterygoid Muscle 3. Occipital Belly of Occipitofrontalis 21. Buccinator Muscle Muscle 22. Cerebrum 4. Auricularis Anterior Muscle 23. Frontal Lobe 5. Auricularis Posterior Muscle 24. Parietal Lobe 6. Auricularis Superior Muscle 25. Occipital Lobe 7. Procerus Muscle 26. Temporal Lobe 8. Orbicularis Oculi Muscle 27. Cerebellum 9. Levator Labii Superioris Alaeque 28. Corpus Callosum Nasi Muscle 29. Septum Pellucidum 10. Levator Labii Superioris Muscle 30. Fornix 11. Zygomaticus Minor Muscle 31. Thalamus 12. Zygomaticus Major Muscle 32. Midbrain 13. Depressor Anguli Oris Muscle 33. Pons 14. Depressor Labii Inferioris Muscle 34. Medulla Oblongata 15. Mentalis Muscle 35. Olfactory Bulb 16. Orbicularis Oris Muscle 36. Optic Nerve II 37. Oculomotor Nerve III II. Body Wall 3. Oblique Fissure 43. Small Cardiac Vein 38. Trochlear Nerve IV 1. Pectoralis Major Muscle 4. Horizontal Fissure of Right Lung 44. Coronary Sinus 39. Trigeminal Nerve V a. Clavicular Head 5. Superior Lobe 40. Abducent Nerve VI b. Sternocostal Head 6. Inferior Lobe IV. Abdominal and Pelvic Viscera 41. Facial Nerve VII c. Abdominal Head 7. Middle Lobe of Right Lung 1. Left Lobe of Liver 42. Vestibulocochlear Nerve VIII 2. Pectoralis Minor Muscle 8. Hilum of Lung 2. Right Lobe of Liver 43. Glossopharyngeal Nerve IX 3. External Intercostal Muscles 9. Trachea 3. Falciform Ligament of Liver 44. Vagus Nerve X 4. Internal Intercostal Muscles 10.