European Jurassic Floras: Statistics and Palaeoenvironmental Proxies
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Pterosaurs Flight in the Age of Dinosaurs Now Open 2 News at the Museum 3
Member Magazine Spring 2014 Vol. 39 No. 2 Pterosaurs Flight in the Age of Dinosaurs now open 2 News at the Museum 3 From the After an unseasonably cold, snowy winter, will work to identify items from your collection, More than 540,000 Marine Fossils the Museum is pleased to offer a number of while also displaying intriguing specimens from President springtime opportunities to awaken the inner the Museum’s own world-renowned collections. Added to Paleontology Collection naturalist in us all. This is the time of year when Of course, fieldwork and collecting have Ellen V. Futter Museum scientists prepare for the summer been hallmarks of the Museum’s work since Collections at a Glance field season as they continue to pursue new the institution’s founding. What has changed, discoveries in their fields. It’s also when Museum however, is technology. With a nod to the many Over nearly 150 years of acquisitions and Members and visitors can learn about their ways that technology is amplifying how scientific fieldwork, the Museum has amassed preeminent own discoveries during the annual Identification investigations are done, this year, ID Day visitors collections that form an irreplaceable record Day in Theodore Roosevelt Memorial Hall. can learn how scientists use digital fabrication of life on Earth. Today, 21st-century tools— Held this year on May 10, Identification Day to aid their research and have a chance to sophisticated imaging techniques, genomic invites visitors to bring their own backyard finds have their own objects scanned and printed on analyses, programs to analyze ever-growing and curios for identification by Museum scientists. -

Cuticle Ultrastructure in Brachyphyllum Garciarum Sp. Nov (Lower Cretaceous, Argentina) Reveals Its Araucarian Affinity
Review of Palaeobotany and Palynology 269 (2019) 104–128 Contents lists available at ScienceDirect Review of Palaeobotany and Palynology journal homepage: www.elsevier.com/locate/revpalbo Cuticle ultrastructure in Brachyphyllum garciarum sp. nov (Lower Cretaceous, Argentina) reveals its araucarian affinity Martin A. Carrizo a,⁎, Maiten A. Lafuente Diaz a, Georgina M. Del Fueyo a, Gaëtan Guignard b a División Paleobotánica, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, CONICET. Av. Ángel Gallardo 470, 1405 Buenos Aires, Argentina b Université Lyon 1, CNRS, UMR 5023 LEHNA, 7-9 rue Raphaël Dubois, Villeurbanne cedex F-69622, Lyon, France article info abstract Article history: A detailed and extensive study of a new species, Brachyphyllum garciarum sp. nov., was carried out through the Received 28 February 2019 analysis of the gross morphology and the cuticle fine details, structure and ultrastructure characters of its leaves Received in revised form 14 June 2019 using light microscope and scanning and transmission electron microscope. The fossils consist of compressions of Accepted 19 June 2019 incomplete twigs with well-preserved cuticle, collected from pelitic levels of the Springhill Formation (lower Hauterivian/lower Barremian) at the Río Correntoso locality in the Santa Cruz province, Argentina. The twigs Keywords: have adpressed scale-like leaves spirally disposed. Leaves have a rhomboidal to pyramidal shape, a width and Foliar cuticle length always in a 1:1 ratio, margin entire and apex mostly rounded. Leaves are amphistomatic with stomatal ap- Ultrastructure paratuses occurring in groups of narrow-wedge shape along the leaf axis. Stomatal apparatuses are close to each Taxonomy other with subsidiary cells in contact; the guard cells are sunken, with marked polar extensions and thickened Brachyphyllum mouth. -

Palynology of the Upper Chinle Formation in Northern New Mexico, U.S.A
Lindström et al. 1 1 Palynology of the upper Chinle Formation in northern New Mexico, U.S.A.: 2 implications for biostratigraphy and terrestrial ecosystem change during the Late 3 Triassic (Norian–Rhaetian) 4 a* b c d 5 Sofie Lindström , Randall B. Irmis , Jessica H. Whiteside , Nathan D. Smith , Sterling J. e f 6 Nesbitt , and Alan H. Turner 7 a 8 Geological Survey of Denmark and Greenland, Øster Voldgade 10, DK-1350 Copenhagen 9 K, DENMARK, [email protected] b 10 Natural History Museum of Utah and Department of Geology & Geophysics, University of 11 Utah, Salt Lake City, UT 84108-1214, USA c 12 Ocean and Earth Science, National Oceanography Centre Southampton, University of 13 Southampton, European Way, Southampton SO14 3ZH, UNITED KINGDOM d 14 Dinosaur Institute, Natural History Museum of Los Angeles County, Los Angeles, CA 15 90007, USA e 16 Department of Geosciences, Virginia Polytechnic Institute and State University, Blacksburg, 17 Virginia 24601 USA f 18 Department of Anatomical Sciences, Stony Brook University, Stony Brook, New York 19 11794-8081, USA 20 21 Abstract 22 A new densely sampled palynological record from the vertebrate-bearing upper Chinle 23 Formation at Ghost Ranch in the Chama Basin of northwestern New Mexico provides insights 24 into the biostratigraphy and terrestrial ecosystem changes during the Late Triassic of 25 northwestern Pangaea. Spore-pollen assemblages from the Poleo Sandstone, Petrified Forest, Lindström et al. 2 26 and 'siltstone' members are dominated by pollen of corystospermous seed ferns (Alisporites) 27 and voltziacean conifers (Enzonalasporites, Patinasporites). Other abundant taxa include 28 Klausipollenites gouldii and the enigmatic fused tetrad Froelichsporites traversei, whereas 29 spores of ferns and fern allies are generally rare. -

Squires Catalogue
Type and Figured Palaeontological Specimens in the Tasmanian Museum and Art Gallery A CATALOGUE Compiled by Tasmanian Museum and Art Gallery Don Squires Hobart, Tasmania Honorary Curator of Palaeontology May, 2012 Type and Figured Palaeontological Specimens in the Tasmanian Museum and Art Gallery A CATALOGUE Compiled by Don Squires Honorary Curator of Palaeontology cover image: Trigonotreta stokesi Koenig 1825, the !rst described Australian fossil taxon occurs abundantly in its type locality in the Tamar Valley, Tasmania as external and internal moulds. The holotype, a wax cast, is housed at the British Museum (Natural History). (Clarke, 1979) Hobart, Tasmania May, 2012 Contents INTRODUCTION ..........................................1 VERTEBRATE PALAEONTOLOGY ...........122 PISCES .................................................. 122 INVERTEBRATE PALAEONTOLOGY ............9 AMPHIBIA .............................................. 123 NEOGENE ....................................................... 9 REPTILIA [SP?] ....................................... 126 MONOTREMATA .................................... 127 PLEISTOCENE ........................................... 9 MARSUPIALIA ........................................ 127 Gastropoda .......................................... 9 INCERTAE SEDIS ................................... 128 Ostracoda ........................................... 10 DESCRIBED AS A VERTEBRATE, MIOCENE ................................................. 14 PROBABLY A PLANT ............................. 129 bivalvia ............................................... -

JUDD W.S. Et. Al. (2002) Plant Systematics: a Phylogenetic Approach. Chapter 7. an Overview of Green
UNCORRECTED PAGE PROOFS An Overview of Green Plant Phylogeny he word plant is commonly used to refer to any auto- trophic eukaryotic organism capable of converting light energy into chemical energy via the process of photosynthe- sis. More specifically, these organisms produce carbohydrates from carbon dioxide and water in the presence of chlorophyll inside of organelles called chloroplasts. Sometimes the term plant is extended to include autotrophic prokaryotic forms, especially the (eu)bacterial lineage known as the cyanobacteria (or blue- green algae). Many traditional botany textbooks even include the fungi, which differ dramatically in being heterotrophic eukaryotic organisms that enzymatically break down living or dead organic material and then absorb the simpler products. Fungi appear to be more closely related to animals, another lineage of heterotrophs characterized by eating other organisms and digesting them inter- nally. In this chapter we first briefly discuss the origin and evolution of several separately evolved plant lineages, both to acquaint you with these important branches of the tree of life and to help put the green plant lineage in broad phylogenetic perspective. We then focus attention on the evolution of green plants, emphasizing sev- eral critical transitions. Specifically, we concentrate on the origins of land plants (embryophytes), of vascular plants (tracheophytes), of 1 UNCORRECTED PAGE PROOFS 2 CHAPTER SEVEN seed plants (spermatophytes), and of flowering plants dons.” In some cases it is possible to abandon such (angiosperms). names entirely, but in others it is tempting to retain Although knowledge of fossil plants is critical to a them, either as common names for certain forms of orga- deep understanding of each of these shifts and some key nization (e.g., the “bryophytic” life cycle), or to refer to a fossils are mentioned, much of our discussion focuses on clade (e.g., applying “gymnosperms” to a hypothesized extant groups. -

A Stable Isotopic Investigation of Resource Partitioning Among Neosauropod Dinosaurs of the Upper Jurassic Morrison Formation
A stable isotopic investigation of resource partitioning among neosauropod dinosaurs of the Upper Jurassic Morrison Formation Benjamin T. Breeden, III SID: 110305422 [email protected] GEOL394H University of Maryland, College Park, Department of Geology 29 April 2011 Advisors: Dr. Thomas R. Holtz1, Jr., Dr. Alan Jay Kaufman1, and Dr. Matthew T. Carrano2 1: University of Maryland, College Park, Department of Geology 2: National Museum of Natural History, Department of Paleobiology ABSTRACT For more than a century, morphological studies have been used to attempt to understand the partitioning of resources in the Morrison Fauna, particularly between members of the two major clades of neosauropod (long-necked, megaherbivorous) dinosaurs: Diplodocidae and Macronaria. While it is generally accepted that most macronarians fed 3-5m above the ground, the feeding habits of diplodocids are somewhat more enigmatic; it is not clear whether diplodocids fed higher or lower than macronarians. While many studies exploring sauropod resource portioning have focused on differences in the morphologies of the two groups, few have utilized geochemical evidence. Stable isotope geochemistry has become an increasingly common and reliable means of investigating paleoecological questions, and due to the resistance of tooth enamel to diagenetic alteration, fossil teeth can provide invaluable paleoecological and behavioral data that would be otherwise unobtainable. Studies in the Ituri Rainforest in the Democratic Republic of the Congo, have shown that stable isotope ratios measured in the teeth of herbivores reflect the heights at which these animals fed in the forest due to isotopic variation in plants with height caused by differences in humidity at the forest floor and the top of the forest exposed to the atmosphere. -

Critical Review of Research on the Lower Jurassic Flora of Poland
Acta Palaeobotanica 53(2): 141–163, 2013 DOI: 10.2478/acpa-2013-0015 Critical review of research on the Lower Jurassic flora of Poland GRZEGORZ PACYNA Department of Palaeobotany and Palaeoherbarium, Institute of Botany, Jagiellonian University, Lubicz 46, 31-512 Kraków, Poland; e-mail: [email protected] Received 7 October 2013; accepted for publication 5 December 2013 ABSTRACT. The Lower Jurassic plant macrofossils of Poland are poorly known. Relatively rich sources of fossils are found in only a few outcrops in the Holy Cross Mountains. Other described plant remains come from drill cores taken from most areas of Poland, but as a rule these are single specimens. The only professional descriptions of Lower Jurassic macroflora are papers by Raciborski, Makarewiczówna, and a team of researchers consisting of Reymanówna, Barbacka, Ziaja, and Wcisło-Luraniec. Raciborski’s fossil collection is still available for research and revision. Such work is in progress. The collection described by Makarewiczówna contained many interesting speci- mens but unfortunately the majority of them are now missing. Stratigraphic research by geologists has provided some new specimens from drill cores and outcrops in the Holy Cross Mountains but these have not been subjected to detailed palaeobotanical analysis. The palynology of the Lower Jurassic was focused on biostratigraphy from the outset of that research. As an outcome it provided spore-pollen and megaspore zonations for Lower Jurassic strata in Poland. The Polish Lower Jurassic flora is comprised of ferns (very numerous), lycopsids, sphenopsids, cycadaleans, bennettitaleans, gnetaleans, ginkgoaleans, and conifers. This flora is taxonomically poorer than the equally old and geographically close floras of Denmark, Sweden, and Germany. -

Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation
i1 Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation William A. DiMichele, William E. Stein, and Richard M. Bateman DiMichele, W.A., Stein, W.E., and Bateman, R.M. 2001. Ecological sorting of vascular plant classes during the Paleozoic evolutionary radiation. In: W.D. Allmon and D.J. Bottjer, eds. Evolutionary Paleoecology: The Ecological Context of Macroevolutionary Change. Columbia University Press, New York. pp. 285-335 THE DISTINCTIVE BODY PLANS of vascular plants (lycopsids, ferns, sphenopsids, seed plants), corresponding roughly to traditional Linnean classes, originated in a radiation that began in the late Middle Devonian and ended in the Early Carboniferous. This relatively brief radiation followed a long period in the Silurian and Early Devonian during wrhich morphological complexity accrued slowly and preceded evolutionary diversifications con- fined within major body-plan themes during the Carboniferous. During the Middle Devonian-Early Carboniferous morphological radiation, the major class-level clades also became differentiated ecologically: Lycopsids were cen- tered in wetlands, seed plants in terra firma environments, sphenopsids in aggradational habitats, and ferns in disturbed environments. The strong con- gruence of phylogenetic pattern, morphological differentiation, and clade- level ecological distributions characterizes plant ecological and evolutionary dynamics throughout much of the late Paleozoic. In this study, we explore the phylogenetic relationships and realized ecomorphospace of reconstructed whole plants (or composite whole plants), representing each of the major body-plan clades, and examine the degree of overlap of these patterns with each other and with patterns of environmental distribution. We conclude that 285 286 EVOLUTIONARY PALEOECOLOGY ecological incumbency was a major factor circumscribing and channeling the course of early diversification events: events that profoundly affected the structure and composition of modern plant communities. -

Fundamentals of Palaeobotany Fundamentals of Palaeobotany
Fundamentals of Palaeobotany Fundamentals of Palaeobotany cuGU .叮 v FimditLU'φL-EjAA ρummmm 吋 eαymGfr 伊拉ddd仇側向iep M d、 況 O C O W Illustrations by the author uc削 ∞叩N Nn凹創 刊,叫MH h 咀 可 白 a aEE-- EEA First published in 1987 by Chapman αndHallLtd 11 New Fetter Lane, London EC4P 4EE Published in the USA by Chα~pman and H all 29 West 35th Street: New Yo地 NY 10001 。 1987 S. V. M秒len Softcover reprint of the hardcover 1st edition 1987 ISBN-13: 978-94-010-7916-7 e-ISBN-13: 978-94-009-3151-0 DO1: 10.1007/978-94-009-3151-0 All rights reserved. No part of this book may be reprinted, or reproduced or utilized in any form or by any electronic, mechanical or other means, now known or hereafter invented, including photocopying and recording, or in any information storage and retrieval system, without permission in writing from the publisher. British Library Cataloguing in Publication Data Mey凹, Sergei V. Fundamentals of palaeobotany. 1. Palaeobotany I. Title 11. Osnovy paleobotaniki. English 561 QE905 Library 01 Congress Catα loging in Publication Data Mey凹, Sergei Viktorovich. Fundamentals of palaeobotany. Bibliography: p. Includes index. 1. Paleobotany. I. Title. QE904.AIM45 561 8ι13000 Contents Foreword page xi Introduction xvii Acknowledgements xx Abbreviations xxi 1. Preservation 抄'pes αnd techniques of study of fossil plants 1 2. Principles of typology and of nomenclature of fossil plants 5 Parataxa and eutaxa S Taxa and characters 8 Peculiarity of the taxonomy and nomenclature of fossil plants 11 The binary (dual) system of fossil plants 12 The reasons for the inflation of generic na,mes 13 The species problem in palaeobotany lS The polytypic concept of the species 17 Assemblage-genera and assemblage-species 17 The cladistic methods 18 3. -
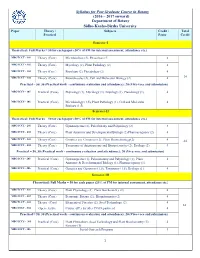
Syllabus for Post Graduate Course in Botany (2016 – 2017 Onward)
Syllabus for Post Graduate Course in Botany (2016 – 2017 onward) Department of Botany Sidho-Kanho-Birsha University Paper Theory / Subjects Credit / Total Practical Paper Credit Semester-I Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 101 Theory (Core) Microbiology (2), Phycology (2) 4 MBOTCCT - 102 Theory (Core) Mycology (2), Plant Pathology (2) 4 MBOTCCT - 103 Theory (Core) Bryology (2), Pteridology (2) 4 MBOTCCT - 104 Theory (Core) Biomolecules (2), Cell and Molecular Biology (2) 4 24 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 105 Practical (Core) Phycology (1), Mycology (1), Bryology (1), Pteridology (1). 4 MBOTCCS - 106 Practical (Core) Microbiology (1.5), Plant Pathology (1), Cell and Molecular 4 Biology (1.5). Semester-II Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 201 Theory (Core) Gymnosperms (2), Paleobotany and Palynology (2) 4 MBOTCCT - 202 Theory (Core) Plant Anatomy and Developmental Biology (2) Pharmacognosy (2) 4 MBOTCCT - 203 Theory (Core) Genetics and Genomics (2), Plant Biotechnology(2) 4 24 MBOTCCT - 204 Theory (Core) Taxonomy of Angiosperms and Biosystematics (2), Ecology (2) 4 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 205 Practical (Core) Gymnosperms (1), Palaeobotany and Palynology (1), Plant 4 Anatomy & Developmental Biology (1), Pharmacognosy (1). MBOTCCS - 206 Practical (Core) Genetics and Genomics (1.5), Taxonomy (1.5), Ecology (1). 4 Semester-III Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 301 Theory (Core) Plant Physiology (2), Plant Biochemistry (2) 4 MBOTCCT - 302 Theory (Core) Economic Botany (2), Bioinformatics (2) 4 MBOTCCT - 303 Theory (Core) Elements of Forestry (2), Seed Technology (2). -

Evolução Das Plantas
EVOLUÇÃO DAS PLANTAS DAS EVOLUÇÃO Esta viagem pela Terra, pela sua formação, pelas suas primeiras atmosferas O segundo volume da coleção «Botânica em e vidas, pela evolução das plantas através das sucessivas mudanças é uma Português» faz uma síntese da história evolutiva leitura fascinante, às vezes difícil, mas como é mostrada e explicada com das plantas, desde a evolução da vida celular nas fontes grande sabedoria transmite conhecimento – saber científico –, que, apesar hidrotermais alcalinas oceânicas, há cerca de quatro EVOLUÇÃO das nossas falhas, conseguimos apreender e aprender. mil milhões de anos, até às grandes florestas tropicais hiperdiversas atuais. Como surgiu a fotossíntese? É uma lição de história, geologia, geografia, climatologia, agronomia As plantas nasceram na água: como invadiram a terra? e biologia, com notas de química e de física, cálculos matemáticos De que modo as plantas interagiram com a atmosfera DAS PLANTAS e paisagísticos, ou seja, a completa Aula de Botânica. terrestre? O que é e qual a origem do solo? Quais as funções das flores, esporos e sementes? Por que razão Carlos Aguiar Neste livro, desde a Pangeia, com a separação dos continentes, até hoje, as plantas com flor são tão bem-sucedidas? De que passando pelas várias erupções, avanços e recuos do mar, degelos e modo as megaextinções influenciaram a evolução das aquecimentos globais, vamos acompanhando os diferentes habitats, a | plantas? Estaremos perante uma nova megaextinção? Carlos Aguiar evolução e transformação das plantas pelos diversos continentes e mares Estas e muitas outras perguntas são respondidas – por seleção natural ou deriva genética – e como se foram aclimatando, ao longo deste livro. -

Fossil and Living Cycads Say No More Megasporophylls
hology orp a Miao et al., J Morphol Anat 2017, 1:2 nd M f A o n l a a t n o r m u y o J Journal of Morphology and Anatomy Research Article Article Open Access Fossil and Living Cycads Say "No More Megasporophylls" Yuyan Miao1,2, Zhong-Jian Liu3, Meina Wang3,4 and Xin Wang5* 1Beijing Museum of Natural History, Beijing, China 2State Key Laboratory of Biogeology and Environmental Geology, China University of Geosciences (Wuhan), Wuhan, China 3Shenzhen Key Laboratory for Orchid Conservation and Utilization, National Orchid Conservation Center of China and Orchid Conservation and Research Center of Shenzhen, Shenzhen, China 4College of Landscape Architecture, Fujian Agriculture and Forestry University, Fuzhou, China 5CAS Key Laboratory of Economic Stratigraphy and Paleogeography, Nanjing Institute of Geology and Palaeontology, Nanjing, China Abstract The origins of angiosperms and cycads are still mysterious. To understand the evolution of these groups as well as other gymnosperms it was impossible without mentioning a frequently used term “megasporophyll”. “Megasporophyll” is a concept that has been used widely in botany. This term is more or less related with the famous saying “Alles ist Blatt” by Goethe. This term became popular since Arber and Parkin hypothesized that the carpels in the Magnoliales were equivalent to and derived from former foliar parts bearing ovules along their margins (“megasporophyll”). Many botanists uncritically called the parts in all the reproductive organs of seed plants as “sporophylls”, no matter what they actually saw in the plants. However, the fact is that none of the reproductive parts (fossil or living), except those in the Cycadales, are foliar or leaf-like.