Pegias, Alasmidonta, and Arcidens
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Penobscot Rivershed with Licensed Dischargers and Critical Salmon
0# North West Branch St John T11 R15 WELS T11 R17 WELS T11 R16 WELS T11 R14 WELS T11 R13 WELS T11 R12 WELS T11 R11 WELS T11 R10 WELS T11 R9 WELS T11 R8 WELS Aroostook River Oxbow Smith Farm DamXW St John River T11 R7 WELS Garfield Plt T11 R4 WELS Chapman Ashland Machias River Stream Carry Brook Chemquasabamticook Stream Squa Pan Stream XW Daaquam River XW Whitney Bk Dam Mars Hill Squa Pan Dam Burntland Stream DamXW Westfield Prestile Stream Presque Isle Stream FRESH WAY, INC Allagash River South Branch Machias River Big Ten Twp T10 R16 WELS T10 R15 WELS T10 R14 WELS T10 R13 WELS T10 R12 WELS T10 R11 WELS T10 R10 WELS T10 R9 WELS T10 R8 WELS 0# MARS HILL UTILITY DISTRICT T10 R3 WELS Water District Resevoir Dam T10 R7 WELS T10 R6 WELS Masardis Squapan Twp XW Mars Hill DamXW Mule Brook Penobscot RiverYosungs Lakeh DamXWed0# Southwest Branch St John Blackwater River West Branch Presque Isle Strea Allagash River North Branch Blackwater River East Branch Presque Isle Strea Blaine Churchill Lake DamXW Southwest Branch St John E Twp XW Robinson Dam Prestile Stream S Otter Brook L Saint Croix Stream Cox Patent E with Licensed Dischargers and W Snare Brook T9 R8 WELS 8 T9 R17 WELS T9 R16 WELS T9 R15 WELS T9 R14 WELS 1 T9 R12 WELS T9 R11 WELS T9 R10 WELS T9 R9 WELS Mooseleuk Stream Oxbow Plt R T9 R13 WELS Houlton Brook T9 R7 WELS Aroostook River T9 R4 WELS T9 R3 WELS 9 Chandler Stream Bridgewater T T9 R5 WELS TD R2 WELS Baker Branch Critical UmScolcus Stream lmon Habitat Overlay South Branch Russell Brook Aikens Brook West Branch Umcolcus Steam LaPomkeag Stream West Branch Umcolcus Stream Tie Camp Brook Soper Brook Beaver Brook Munsungan Stream S L T8 R18 WELS T8 R17 WELS T8 R16 WELS T8 R15 WELS T8 R14 WELS Eagle Lake Twp T8 R10 WELS East Branch Howe Brook E Soper Mountain Twp T8 R11 WELS T8 R9 WELS T8 R8 WELS Bloody Brook Saint Croix Stream North Branch Meduxnekeag River W 9 Turner Brook Allagash Stream Millinocket Stream T8 R7 WELS T8 R6 WELS T8 R5 WELS Saint Croix Twp T8 R3 WELS 1 Monticello R Desolation Brook 8 St Francis Brook TC R2 WELS MONTICELLO HOUSING CORP. -

Francis Marion National Forest Freshwater Mussel Surveys: Final Report
Francis Marion National Forest Freshwater Mussel Surveys: Final Report Contract No. AG-4670-C-10-0077 Prepared For: Francis Marion and Sumter National Forests 4931 Broad River Road Columbia, SC 29212-3530 Prepared by: The Catena Group, Inc. 410-B Millstone Drive Hillsborough, NC 27278 November 2011 TABLE OF CONTENTS 1.0 INTRODUCTION ................................................................................................... 1 1.1 Background and Objectives ................................................................................. 1 2.0 STUDY AREA ........................................................................................................ 2 3.0 METHODS .............................................................................................................. 2 4.0 RESULTS ................................................................................................................ 3 5.0 DISCUSSION ........................................................................................................ 19 5.1 Mussel Habitat Conditions in the FMNF (Objective 1) ..................................... 19 5.2 Mussel Fauna of the FMNF (Objective 2) ......................................................... 19 5.2.1 Mussel Species Found During the Surveys ................................................ 19 5.3 Highest Priority Mussel Fauna Streams of the FMNF (Objective 3) ................. 25 5.4 Identified Areas that Warrant Further Study in the FMNF (Objective 4) .......... 25 6.0 LITERATURE CITED ......................................................................................... -
Field Identification Guide to the Living Marine Resources In
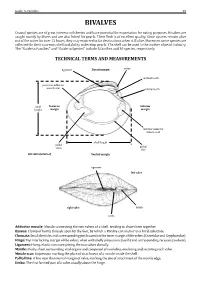
Guide to Families 29 BIVALVES Coastal species are of great interest to fisheries and have potential for exportation for eating purposes. Bivalves are caught mainly by divers and are also fished for pearls. Their flesh is of excellent quality. Since oysters remain alive out of the water for over 12 hours, they may exported to far destinations when still alive. Moreover, some species are collected for their nacreous shell and ability to develop pearls. The shell can be used in the mother of pearl industry. The “Guide to Families’’ andTECHNICAL ‘‘Guide to Species’’ TERMS include 5AND families MEASUREMENTS and 10 species, respectively. ligament Dorsal margin umbo posterior adductor cardinal tooth muscle scar lateral tooth Posterior Anterior margin margin shell height anterior adductor muscle scar pallial sinus pallial shell length line left valve (interior) Ventral margin ligament left valve right valve lunule umbo Adductor muscle: Byssus: Chomata: Muscle connecting the two valves of a shell, tending to draw them together. Hinge: Clump of horny threads spun by the foot, by which a Bivalve can anchor to a hard substrate. Ligament: Small denticles and corresponding pits located on the inner margin of the valves (Ostreidae and Gryphaeidae). Mantle: Top interlocking margin of the valves, often with shelly projections (teeth) and corresponding recesses (sockets). Muscle scar: Horny, elastic structure joining the two valves dorsally. Pallial line: Fleshy sheet surrounding vital organs and composed of two lobes, one lining and secreting each valve. Umbo: Impression marking the place of attachment of a muscle inside the shell. A line near the internal margin of valve, marking the site of attachment of the mantle edge. -

Tennessee's Extinct Species
Tennessee's Extinct Species The following species Birds: once occurred in Carolina parakeet Conuropsis carolinensis Ectopistes migratorius Tennessee and are now Passenger pigeon believed to be extinct. Mammals: Following this list are two Eastern elk species descriptions-one Fishes: describing the Carolina Harelip sucker parakeet and another describing the extinct Mussels: Acornshell Epioblasma haysiana freshwater mussels Angled riffleshell Epioblasma biemarginata of Tennessee. Cumberland leafshell Epioblasma stewardsoni Leafshell Epioblasma flexuosa Narrowcat's paw Epioblasma lenoir Rough rockshell Quadrula tuberosa Round combshell Epioblasma personata Sugarspoon Epioblasma arcaeformis Tennessee riffleshell Epioblasma propinqua Carolina Parakeet Status Habitat The Carolina parakeet is an The Carolina parakeet was found Learn rrwreabout extinct species. in riverine forests, cypress swamps, Tennessee's diverse and other woodlands over much of Description the Eastern and Midwest Regions of ecosyster.n3.Su~ort The Carolina parakeet was a the United States. It was the only conservation in your small parrot, about 12inches in parrot native to the United States. community and state! length. Its head was lemon yellow, The parakeets rested at night in with an orange forehead and cheeks. groups, with as many as 30 birds The rest of its body was green. Its sleeping inside one hollowtree, while legs and beak were pale pinkish- others would hang on the outside. white. These curious birds lived and Nests were placed in hollowtrees, traveled in flocks. and three to five white eggs were laid. Up to 50 nests were often crowded into one tree. Role in the Ecosystem Carolina parakeets enjoyed a variety of different foods-apples, peaches, mulberries, pecans, grapes, dogwood fruit, and grains. -

2012 Annual Report Maine Public Reserved
2012 ANNUAL REPORT to the JOINT STANDING COMMITTEE on AGRICULTURE, CONSERVATION AND FORESTRY MAINE PUBLIC RESERVED, NONRESERVED, AND SUBMERGED LANDS Seboeis Lake Unit MAINE DEPARTMENT OF AGRICULTURE, CONSERVATION AND FORESTRY Division of Parks and Public Lands March 1, 2013 2012 ANNUAL REPORT Maine Department of Agriculture, Conservation and Forestry Division of Parks and Public Lands TABLE OF CONTENTS I. INTRODUCTION…………………………………………………………………….. 1 II. 2012 HIGHLIGHTS…………………………………………………………………..1 III. SCOPE OF RESPONSIBILITIES………………………………………………….. 4 IV. LAND MANAGEMENT PLANNING……………………………………………….. 8 V. NATURAL/HISTORIC RESOURCES……………………………………………...9 VI. WILDLIFE RESOURCES……………………………………………………………11 VII. RECREATION RESOURCES………………………………………………………12 VIII. TIMBER RESOURCES……………………………………………………………...18 IX. TRANSPORTATION………………………………………………………………… 22 X. PUBLIC ACCESS…………………………………………………………………… 22 XI. LAND TRANSACTIONS……………………………………………………………. 23 XII. SUBMERGED LANDS……………………………………………………………… 24 XIII. SHORE AND HARBOR MANAGEMENT FUND………………………………… 24 XIV. COASTAL ISLAND PROGRAM…………………………………………………… 24 XV. ADMINISTRATION………………………………………………………………….. 25 XVI. INCOME AND EXPENDITURES ACCOUNTING – CY 2012………………….. 26 XVII. FINANCIAL REPORT FOR FY 2014……………………………………………… 29 XVIII. CONCLUSION……………………………………………………………………….. 36 Appendix A: Map of DPPL Properties & Acquisitions in 2012…………………. 37 Appendix B: Table of 2012 DPPL Acquisitions and Dispositions………………… 38 Appendix C: Federal Forest Legacy Projects Approved and Pending……………... 39 Cover Photo by Kathy Eickenberg -

THE NAUTILUS (Quarterly)
americanmalacologists, inc. PUBLISHERS OF DISTINCTIVE BOOKS ON MOLLUSKS THE NAUTILUS (Quarterly) MONOGRAPHS OF MARINE MOLLUSCA STANDARD CATALOG OF SHELLS INDEXES TO THE NAUTILUS {Geographical, vols 1-90; Scientific Names, vols 61-90) REGISTER OF AMERICAN MALACOLOGISTS JANUARY 30, 1984 THE NAUTILUS ISSN 0028-1344 Vol. 98 No. 1 A quarterly devoted to malacology and the interests of conchologists Founded 1889 by Henry A. Pilsbry. Continued by H. Burrington Baker. Editor-in-Chief: R. Tucker Abbott EDITORIAL COMMITTEE CONSULTING EDITORS Dr. William J. Clench Dr. Donald R. Moore Curator Emeritus Division of Marine Geology Museum of Comparative Zoology School of Marine and Atmospheric Science Cambridge, MA 02138 10 Rickenbacker Causeway Miami, FL 33149 Dr. William K. Emerson Department of Living Invertebrates Dr. Joseph Rosewater The American Museum of Natural History Division of Mollusks New York, NY 10024 U.S. National Museum Washington, D.C. 20560 Dr. M. G. Harasewych 363 Crescendo Way Dr. G. Alan Solem Silver Spring, MD 20901 Department of Invertebrates Field Museum of Natural History Dr. Aurele La Rocque Chicago, IL 60605 Department of Geology The Ohio State University Dr. David H. Stansbery Columbus, OH 43210 Museum of Zoology The Ohio State University Dr. James H. McLean Columbus, OH 43210 Los Angeles County Museum of Natural History 900 Exposition Boulevard Dr. Ruth D. Turner Los Angeles, CA 90007 Department of Mollusks Museum of Comparative Zoology Dr. Arthur S. Merrill Cambridge, MA 02138 c/o Department of Mollusks Museum of Comparative Zoology Dr. Gilbert L. Voss Cambridge, MA 02138 Division of Biology School of Marine and Atmospheric Science 10 Rickenbacker Causeway Miami, FL 33149 EDITOR-IN-CHIEF The Nautilus (USPS 374-980) ISSN 0028-1344 Dr. -

Status and Population Genetics of the Alabama Spike (Elliptio Arca) in the Mobile River Basin
STATUS AND POPULATION GENETICS OF THE ALABAMA SPIKE (ELLIPTIO ARCA) IN THE MOBILE RIVER BASIN A Thesis by DANIEL HUNT MASON Submitted to the Graduate School at Appalachian State University in partial fulfillment of the requirements for the degree of MASTER OF SCIENCE August, 2017 Department of Biology STATUS AND POPULATION GENTICS OF THE ALABAMA SPIKE (ELLIPTIO ARCA) IN THE MOBILE RIVER BASIN A Thesis by DANIEL HUNT MASON August, 2017 APPROVED BY: Michael M. Gangloff, Ph.D. Chairperson, Thesis Committee Matthew C. Estep, Ph.D. Member, Thesis Committee Lynn M. Siefermann, Ph.D. Member, Thesis Committee Zack E. Murrell, Ph.D. Chairperson, Department of Biology Max C. Poole, Ph.D. Dean, Cratis D. Williams School of Graduate Studies Copyright by Daniel Hunt Mason 2017 All Rights Reserved Abstract STATUS AND POPULATION GENETICS OF THE ALABAMA SPIKE (ELLIPTIO ARCA) IN THE MOBILE RIVER BASIN Daniel H. Mason B.A., Appalachian State University M.A., Appalachian State University Chairperson: Dr. Michael M. Gangloff Declines in freshwater mussels (Bivalvia: Unionioda) are widely reported but rarely rigorously tested. Additionally, the population genetics of most species are virtually unknown, despite the importance of these data when assessing the conservation status of and recovery strategies for imperiled mussels. Freshwater mussel endemism is high in the Mobile River Basin (MRB) and many range- restricted taxa have been heavily impacted by riverine alterations, and many species are suspected to be declining in abundance, including the Alabama Spike (Elliptio arca). I compiled historical and current distributional data from all major MRB drainages to quantify the extent of declines in E. -

Final Report- HWY-2009-16 Propagation and Culture of Federally Listed Freshwater Mussel Species
Final Report- HWY-2009-16 Propagation and Culture of Federally Listed Freshwater Mussel Species Prepared By Jay F- Levine, Co-Principal Investigator1 Christopher B- Eads, Co-Investigator1 Renae Greiner, Graduate Student Assistant1 Arthur E- Bogan, Co- Investigator2 1North Carolina State University College of Veterinary Medicine 4700 Hillsborough Street Raleigh, NC 27606 2 NC State Museum of Natural Sciences 4301 Reedy Creek Rd- Raleigh, NC 27607 November 2011 Technical Report Documentation Page 1- Report No- 2-Government Accession No- 3- Recipient’s Catalog No- FHWA/NC/2009-16 4- Title and Subtitle 5- Report Date Propagation and Culture of Federally Listed Freshwater November 2011 Mussel Species 6-Performing Organization Code 7- Author(s) 8-Performing Organization Report No- Jay F- Levine, Co-Principal Investigator Arthur E- Bogan, Co-Principal Investigator Renae Greiner, Graduate Student Assistant 9- Performing Organization Name and Address 10- Work Unit No- (TRAIS) North Carolina State University College of Veterinary Medicine 11- Contract or Grant No- 4700 Hillsborough Street Raleigh, NC 27606 12- Sponsoring Agency Name and Address 13-Type of Report and Period Covered North Carolina Department of Transportation Final Report P-O- Box 25201 August 16, 2008 – June 30, 2011 Raleigh, NC 27611 14- Sponsoring Agency Code HWY-2009-16 15- Supplementary Notes 16- Abstract Road and related crossing construction can markedly alter stream habitat and adversely affect resident native flora. The National Native Mussel Conservation Committee has recognized artificial propagation and culture as an important potential management tool for sustaining remaining freshwater mussel populations and has called for additional propagation research to help conserve and restore this faunal group. -

FAO Fisheries & Aquaculture
Food and Agriculture Organization of the United Nations Fisheries and for a world without hunger Aquaculture Department Cultured Aquatic Species Information Programme Mercenaria mercenaria (Linnaeus, 1758) I. Identity V. Status And Trends a. Biological Features VI. Main Issues b. Images Gallery a. Responsible Aquaculture Practices II. Profile VII. References a. Historical Background a. Related Links b. Main Producer Countries c. Habitat And Biology III. Production a. Production Cycle b. Production Systems c. Diseases And Control Measures IV. Statistics a. Production Statistics b. Market And Trade Identity Mercenaria mercenaria Linnaeus, 1758 [Veneridae] FAO Names: En - Northern quahog(=Hard clam), Fr - Praire, Es - Chirla mercenaria Biological features Shell solid, equivalve; inequilateral, beaks in the front half of the shell; broadly oval in outline. Ligament a deeply inset, dark brown elliptical band, behind the beaks reaching half-way to the posterior margin. Lunule well defined, broad, heart-shaped. Escutcheon indistinct. Sculpture of concentric lines, raised here and there into ridges, and fine radiating lines. In young specimens the ridges are present all over the shell but in the adult they persist, after wear and tear, only near the anterior and posterior margins. Growth stages prominent. Both valves with three cardinal teeth; in addition there is present in each valve a rough tooth-like area behind the beaks and immediately below the ligament; this area has the appearance of a supplementary posterior cardinal tooth which has been broken off. No laterals. Pallial sinus not deep, triangular. Margin grenulate. Colour a dirty white, light varnish-brown, dull grey or grey-brown. Inside of shell white, sometimes deep violet about the adductor muscle scars. -

Restoring the Endangered Oyster Mussel (Epioblasma Capsaeformis) to the Upper Clinch River, Virginia: an Evaluation of Population Restoration Techniques Caitlin S
RESEARCH ARTICLE Restoring the endangered oyster mussel (Epioblasma capsaeformis) to the upper Clinch River, Virginia: an evaluation of population restoration techniques Caitlin S. Carey1,2,3,JessW.Jones4, Robert S. Butler5, Eric M. Hallerman6 From 2005 to 2011, the federally endangered freshwater mussel Epioblasma capsaeformis (oyster mussel) was reintroduced at three sites in the upper Clinch River, Virginia, using four release techniques. These release techniques were (1) translocation of adults (site 1, n = 1418), (2) release of laboratory-propagated sub-adults (site 1, n = 2851), (3) release of 8-week-old laboratory-propagated juveniles (site 2, n = 9501), and (4) release of artificially infested host fishes (site 3, n = 1116 host fishes). These restoration efforts provided a unique research opportunity to compare the effectiveness of techniques used to reestablish populations of extirpated and declining species. We evaluated the relative success of these four population restoration approaches via monitoring at each release site (2011–2012) using systematic 0.25-m2 quadrat sampling to estimate abundance and post-release survival. Abundances of translocated adult and laboratory-propagated sub-adult E. capsaeformis at site 1 ranged 577–645 and 1678–1700 individuals, respectively, signifying successful settlement and high post-release survival. Two untagged individuals (29.1 and 27.3 mm) were observed, indicating that recruitment is occurring at site 1. No E. capsaeformis were found at sites where 8-week-old laboratory-propagated juveniles (site 2) and artificially infested host fishes (site 3) were released. Our results indicate that translocations of adults and releases of laboratory-propagated sub-adults were the most effective population restoration techniques for E. -

Guiding Species Recovery Through Assessment of Spatial And
Guiding Species Recovery through Assessment of Spatial and Temporal Population Genetic Structure of Two Critically Endangered Freshwater Mussel Species (Bivalvia: Unionidae) Jess Walter Jones ( [email protected] ) United States Fish and Wildlife Service Timothy W. Lane Virginia Department of Game and Inland Fisheries N J Eric M. Hallerman Virginia Tech: Virginia Polytechnic Institute and State University Research Article Keywords: Freshwater mussels, Epioblasma brevidens, E. capsaeformis, endangered species, spatial and temporal genetic variation, effective population size, species recovery planning, conservation genetics Posted Date: March 16th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-282423/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/28 Abstract The Cumberlandian Combshell (Epioblasma brevidens) and Oyster Mussel (E. capsaeformis) are critically endangered freshwater mussel species native to the Tennessee and Cumberland River drainages, major tributaries of the Ohio River in the eastern United States. The Clinch River in northeastern Tennessee (TN) and southwestern Virginia (VA) harbors the only remaining stronghold population for either species, containing tens of thousands of individuals per species; however, a few smaller populations are still extant in other rivers. We collected and analyzed genetic data to assist with population restoration and recovery planning for both species. We used an 888 base-pair sequence of the mitochondrial NADH dehydrogenase 1 (ND1) gene and ten nuclear DNA microsatellite loci to assess patterns of genetic differentiation and diversity in populations at small and large spatial scales, and at a 9-year (2004 to 2013) temporal scale, which showed how quickly these populations can diverge from each other in a short time period. -

Atlas of the Freshwater Mussels (Unionidae)
1 Atlas of the Freshwater Mussels (Unionidae) (Class Bivalvia: Order Unionoida) Recorded at the Old Woman Creek National Estuarine Research Reserve & State Nature Preserve, Ohio and surrounding watersheds by Robert A. Krebs Department of Biological, Geological and Environmental Sciences Cleveland State University Cleveland, Ohio, USA 44115 September 2015 (Revised from 2009) 2 Atlas of the Freshwater Mussels (Unionidae) (Class Bivalvia: Order Unionoida) Recorded at the Old Woman Creek National Estuarine Research Reserve & State Nature Preserve, Ohio, and surrounding watersheds Acknowledgements I thank Dr. David Klarer for providing the stimulus for this project and Kristin Arend for a thorough review of the present revision. The Old Woman Creek National Estuarine Research Reserve provided housing and some equipment for local surveys while research support was provided by a Research Experiences for Undergraduates award from NSF (DBI 0243878) to B. Michael Walton, by an NOAA fellowship (NA07NOS4200018), and by an EFFRD award from Cleveland State University. Numerous students were instrumental in different aspects of the surveys: Mark Lyons, Trevor Prescott, Erin Steiner, Cal Borden, Louie Rundo, and John Hook. Specimens were collected under Ohio Scientific Collecting Permits 194 (2006), 141 (2007), and 11-101 (2008). The Old Woman Creek National Estuarine Research Reserve in Ohio is part of the National Estuarine Research Reserve System (NERRS), established by section 315 of the Coastal Zone Management Act, as amended. Additional information on these preserves and programs is available from the Estuarine Reserves Division, Office for Coastal Management, National Oceanic and Atmospheric Administration, U. S. Department of Commerce, 1305 East West Highway, Silver Spring, MD 20910.