Final Report of the Eastern Slopes Grizzly Bear Project
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Highway 3: Transportation Mitigation for Wildlife and Connectivity in the Crown of the Continent Ecosystem
Highway 3: Transportation Mitigation for Wildlife and Connectivity May 2010 Prepared with the: support of: Galvin Family Fund Kayak Foundation HIGHWAY 3: TRANSPORTATION MITIGATION FOR WILDLIFE AND CONNECTIVITY IN THE CROWN OF THE CONTINENT ECOSYSTEM Final Report May 2010 Prepared by: Anthony Clevenger, PhD Western Transportation Institute, Montana State University Clayton Apps, PhD, Aspen Wildlife Research Tracy Lee, MSc, Miistakis Institute, University of Calgary Mike Quinn, PhD, Miistakis Institute, University of Calgary Dale Paton, Graduate Student, University of Calgary Dave Poulton, LLB, LLM, Yellowstone to Yukon Conservation Initiative Robert Ament, M Sc, Western Transportation Institute, Montana State University TABLE OF CONTENTS List of Tables .....................................................................................................................................................iv List of Figures.....................................................................................................................................................v Executive Summary .........................................................................................................................................vi Introduction........................................................................................................................................................1 Background........................................................................................................................................................3 -
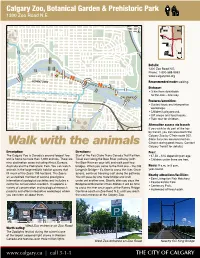
Calgary Zoo Commute
Calgary Zoo, Botanical Garden & Prehistoric Park 1300 Zoo Road N.E. ± NW NE TRANS CENTRE ST CANADA BO W HWY RI VER MEMORIAL DR TR MACLEOD SW SE 2" Details: 1300 Zoo Road N.E. ! ! ! Phone: 1-800-588-9993 ! ! ! 1 www.calgaryzoo.org ! .1 ! ! ! DOWNTOWN ! ! ! ! ! Recommended mode: walking. ! ! ! 1.3 ! ! ! 2" ! ! ! ! Distance: ! ! 2" 2" 2" 2" 2" • 3 km from downtown 2" 2" 2" ! 2" 2" 2" 2" to the Zoo – one way. CALGARY ZOO Features/amenities: • Guided tours and interpretive workshops. • Children’s playground. • Gift shops and food kiosks. 2" • Train tour for children. Alternative access via transit: If you wish to do part of the trip by transit, you can also reach the Calgary Zoo by CTrain route 202. (Note: bicycles are restricted on Walk with the animals CTrains during peak hours. Contact Calgary Transit for details.) Description: Directions: Fees: The Calgary Zoo is Canada’s second largest zoo Start at the Eau Claire Trans Canada Trail Pavilion. • $7.50 – $16 depending on age. and is home to more than 1,000 animals. There are Travel east along the Bow River pathway (with • Children under three are free. nine destination areas including Africa, Eurasia, the Bow River on your left) and walk past two Australia and the Prehistoric Park. You can watch bridges. When you come to the third one – the Old Hours: 9 a.m. to 5 p.m. animals in the large realistic habitat spaces that Langevin Bridge – it’s time to cross the river. Once year-round. fill much of the Zoo’s 159 hectares. -

REGULAR MEETING of COUNCIL Town Hall, 305 Iles Way
REGULAR MEETING OF COUNCIL Town Hall, 305 Iles Way MISSION STATEMENT Council is committed to excellence in government that reflects our citizens’ interests, values and goals. We will strive to deliver quality public services in a proactive and responsible manner while ensuring our objectives are clearly communicated to our citizens. We will strive to strengthen and diversify our economy by balancing public safety, industrial development, environmental responsibility and sustainability. MONDAY, JULY 13, 2020 5:00 PM A G E N D A Page 1. CALL TO ORDER 2. APPROVAL OF AGENDA 2.1 APPROVAL OF AGENDA RECOMMENDATION THAT the Monday, July 13, 2020 Regular Council Meeting agenda be adopted as presented. 3. ADOPTION OF MINUTES 9 - 11 3.1 JUNE 8, 2020 SPECIAL COUNCIL MEETING MINUTES RECOMMENDATION THAT the minutes from the Monday, June 8, 2020 Special Meeting of Council Page 1 of 139 be adopted as presented. Special Meeting of Council - 08 Jun 2020 - Minutes - Pdf 12 - 19 3.2 JUNE 15, 2020 REGULAR COUNCIL MEETING MINUTES RECOMMENDATION THAT the minutes from the Monday, June 15, 2020 Regular Meeting of Council be adopted as presented. Regular Meeting of Council - 15 Jun 2020 - Minutes - Pdf 20 - 21 3.3 JUNE 29, 2020 SPECIAL COUNCIL MEETING RECOMMENDATION THAT the minutes from the Monday, June 29, 2020 Special Meeting of Council be adopted as presented. Special Meeting of Council - 29 Jun 2020 - Minutes - Pdf 4. BUSINESS ARISING FROM THE MINUTES 5. PROCLAMATIONS None 6. DELEGATIONS AND PETITIONS None 7. CONSENT AGENDA 7.1 CONSENT AGENDA RECOMMENDATION THAT all items in the Monday, July 13, 2020 consent agenda be moved for information. -

Southern Continental Divide Goat 2009...12
WMU 402 Goat Survey – ACA/ASRD D‐AUS Report July 2009 By Alberta Conservation Association And Alberta Sustainable Resource Development, Fish and Wildlife July, 2009 Blairmore, Alberta PERMISSION TO QUOTE This report contains preliminary information and interpretations and may be subject to future revision. To prevent the issuance of misleading information, persons wishing to quote from this report, to cite it in bibliographies or to use it in any other form must first obtain permission from the Executive Director of the Wildlife Management Branch, Fish and Wildlife Division and / or its regional representatives within the Southern Rockies Area. ACKNOWLEDGEMENTS The 2009 WMU 402 goat survey was navigated by Greg Hale (FW), while Mike Jokinen (ACA), Jon Jorgenson (FW) and Rob Watt (Parks Canada‐Waterton) participated as observers. The aircraft was chartered from Bighorn Helicopters with Greg Goodison as the pilot. ABSTRACT An aerial survey to monitor the status of the mountain goat population in Wildlife Management Unit (WMU) 402 was conducted on July 11th, 12th and 15th, 2009. Survey objectives included the collection of herd distribution, herd composition and total goat numbers. All mountain complexes associated with the WMU 402 goat survey received detailed aerial coverage during the survey. The WMU 402 survey area is the northern portion of Goat Management Area (GMA)–A, and currently does not have a designated goat hunting area. A total of 186 mountain goats (119 adults, 44 kids and 23 yearlings) were observed during the survey. Kid and yearling ratios per 100 adults were 37 and 19 respectively. Sixty nine goats were observed on the Alberta side of the Continental Divide while 41 goats were observed in British Columbia. -

Learning with Wetlands at the Sam Livingston Fish Hatchery: a Marriage of Mind and Nature
University of Calgary PRISM: University of Calgary's Digital Repository Graduate Studies Legacy Theses 1999 Learning with wetlands at the Sam Livingston fish hatchery: A marriage of mind and nature Grieef, Patricia Lynn Grieef, P. L. (1999). Learning with wetlands at the Sam Livingston fish hatchery: A marriage of mind and nature (Unpublished master's thesis). University of Calgary, Calgary, AB. doi:10.11575/PRISM/12963 http://hdl.handle.net/1880/25035 master thesis University of Calgary graduate students retain copyright ownership and moral rights for their thesis. You may use this material in any way that is permitted by the Copyright Act or through licensing that has been assigned to the document. For uses that are not allowable under copyright legislation or licensing, you are required to seek permission. Downloaded from PRISM: https://prism.ucalgary.ca The University of Calgary Leurnhg with wetiads at the Sam Livingston Fish Hatchery: A Marriage of Mind and Nature by Patricia L. Grieef A Master's Degree Project submitted to the Faculty of Environmental Design in partial hlfillment of the requirements for the degree of Master of Environmental Design (Environmental Science) Calgary, Alberta September, 1999 O Patricia L. Grieef, 1999 National Library BibliotWque nationale 1*1 .,&"a& du Canada Acquisitions and Acquisitions et Bibliographic Services services bibliographiques 395 Wellington Street 395. nn, Wellington OttawaON KlAW OCtewaON K1AON4 Canada Canada The author has granted a non- L'auteur a accorde une licence non exclusive licence allowing the exclusive pennettant a la National Library of Canada to Bibliotheque nationale du Canada de reproduce, loan, distribute or sell reproduire, preter, distribuer ou copies of this thesis in microform, vendre des copies de cette these sous paper or electronic formats. -

District of Elkford: Climate Change Adaptation Strategy
District of Elkford: Climate Change Adaptation Strategy Report for : District of Elkford, B.C., Canada Developed by: Karen Gorecki BSc, MPP Megan Walsh, BSc Env, MSc Env Mgmt. (2010) and Jeff Zukiwsky BTM, MRM Planning (2009) Executive Summary For several years, the Columbia Basin Trust (CBT) has been spearheading research on climate impacts in the Columbia Basin, supported by a team of scientists and planners, and in partnership with the Pacific Climate Impacts Consortium (PCIC). The CBT observed that the impacts of climate change, and the vulnerability of communities to change, varied throughout the Basin. In 2008, the CBT recognized the need to support communities in identifying localized climate change projections and vulnerabilities, and in creating adaptation plans that would allow those communities to prepare for a much different future. The District of Elkford was one of two communities chosen to participate in Columbia Basin Trust’s Communities Adapting to Climate Change (CACC) initiative. For the District of Elkford, this initiative was integrated into an Official Community Plan (OCP) revision, which was to be completed ‘through a climate change lens.’ With support from PCIC, the Communities Adapting to Climate Change team (comprised of Karen Gorecki, Megan Walsh and Jeff Zukiwsky) developed a process that emphasized community and staff engagement, and the pairing of local knowledge with scientific data and projections to determine areas of priority for further climate impact research. Initial open houses revealed strong community concern for issues such as wildfire, road maintenance, stormwater management and water quality. The CACC research team used the public input and best available science to focus in on three priority areas: Wildfire, Flooding and Water Supply. -

Elk Valley Transit Sparwood and Fernie
Holiday Service Welcome Aboard Fares all fares subject to change Your local transit system runs five days a week. Service is not available on Saturday, Sunday or the Buses serve the Elk Valley, connecting Elkford, following holidays: Cash Elk Valley Transit Sparwood and Fernie. • New Years Day • B.C. Day Adult/Student/Senior $ 2.00 Fixed-route service – scheduled service to major • Family Day • Labour Day Child, 4 or under free destinations and residential areas. • Good Friday • Thanksgiving Day • Easter Monday • Remembrance Day Tickets (10) Health Connections – booked service to Cranbrook. RIDER’S GUIDE • Victoria Day • Christmas Day Adult/Student/Senior 15.00 About Your Transit System • Canada Day • Boxing Day Effective January 2, 2013 Monthly Pass Funding for your local transit system is cost shared Updated July 2020 Subject to change. Check online at bctransit.com for Adult/Student/Senior 42.00 between the Regional District of East Kootenay and special event service. BC Transit. Health Connections service is administered by the Kootenay East Regional Hospital District and Ticket and Pass Outlets BC Transit and funded by the Interior Health Authority. Pass Programs Cranbrook: Regional District of East Kootenay Office Decisions on fares, routes and service levels are made Transit Pass Tax Credit Elkford: District Office by the Regional District of East Kootenay board based The Government of Canada offers a non-refundable Fernie: City Hall on public feedback and information provided by tax credit if you use a monthly pass. Save your pass Sparwood: Leisure Centre BC Transit. Decisions on Health Connections service are and receipt as proof of purchase. -

AGENDA November 14, 2017 5:30 P.M
DISTRICT OF ELKFORD COMMITTEE OF THE WHOLE AGENDA November 14, 2017 5:30 P.M. Council Chambers Elkford's Mission - Through innovative leadership we provide opportunities for responsible growth, in harmony with industry and the environment. We take advantage of opportunities that enhance affordable community living and sustain the quality of life citizens, businesses and visitors expect. Page 1 APPROVAL OF AGENDA (a) Approval of November 14, 2017 Agenda 2 ADOPTION OF MINUTES 3 - 4 (a) Adoption of August 28, 2017 Minutes 3 DELEGATIONS 5 - 134 (a) Draft Community Wildfire Protection Plan • Presenter: Tove Pashkowski, B.A. Blackwell & Associates Ltd. 4 ADJOURNMENT (a) Move to Adjourn Page 1 of 134 Page 2 of 134 DISTRICT OF ELKFORD COMMITTEE OF THE WHOLE Minutes August 28, 2017 Present: Mayor McKerracher, Chair Councillor McGregor Councillor Fairbairn Councillor Wildeman Councillor Bertrand Councillor Zarowny Absent: Councillor Robinson Staff Present: Curtis Helgesen, Chief Administrative Officer Scott Beeching, Director, Planning and Development Services Garity Stanley, Director, Leisure Services Duane Allen, Superintendent, Public Works Marilyn Rookes, Director, Financial Services Corey Kortmeyer, Director, Fire Rescue and Emergency Services Curtis Nyuli, Deputy Director, Fire Rescue and Emergency Services Dorothy Szawlowski, Deputy Director, Corporate Services, Recorder There being a quorum of Council, Mayor McKerracher called the meeting to order at 5:37 pm. APPROVAL OF AGENDA (a) Approval of August 28, 2017 Agenda Moved, Seconded AND RESOLVED THAT the agenda for the August 28, 2017 Committee of the Whole Meeting be approved as circulated. CARRIED ADOPTION OF MINUTES (a) Adoption of August 14, 2017 Minutes Moved, Seconded AND RESOLVED THAT the minutes from the August 14, 2017 Committee of the Whole Meeting be adopted as circulated. -
Banff National Park Offers Many More Helen Katherine Backcountry Opportunities Than Those Lake Lake PARK Trail Shelters Berry River Described Here
BACKCOUNTRY CAMPGROUNDS JASPER CAMPGR OUND TOPO MAP NO . GRID REF . CAMPGR OUND TOPO MAP NO . GRID REF . WHITE GOAT NATIONAL Nigel Ba15 Wildflower Creek 82 N/8 686-003 * Lm20 Mount Costigan 82 0/3 187-783 Pass Bo1c Bow River/canoe 82 0/4 802-771 * Lm22 The Narrows 82 0/6 200-790 PARK * Br9 Big Springs 82 J/14 072-367 Lm31 Ghost Lakes 82 0/6 210-789 Sunwapta WILDERNESS AREA ◊ Br13 Marvel Lake 82 J/13 043-387 ◊ Ml22 Mystic Valley 82 0/5 886-824 Mount Pass Abraham Snowdome Lake Br14 McBride’s Camp 82 J/13 041-396 Mo5 Mosquito Creek 82 N/9 483-240 Mount Br17 Allenby Junction 82 J/13 016-414 * Mo16 Molar Creek 82 N/9 555-154 BIA Athabasca * Bw10 Brewster Creek 82 0/4 944-600 ◊ Mo18 Fish Lakes 82 N/9 556-217 NORTH * Cr6 Cascade Bridge 82 0/5 022-827 * No5 Norman Lake 83 C/2 071-706 * Cr15 Stony Creek 82 0/5 978-896 ◊ Pa8 Paradise Valley 82 N/8 528-898 * Cr31 Flints Park 82 0/5 862-958 * Re6 Lost Horse Creek 82 0/4 784-714 COLUM Glacier 93 Saskatchewan * Cr37 Block Lakes Junction 82 0/5 815-935 Re14 Shadow Lake 82 0/4 743-691 Cs Castleguard 82 C/3 857-703 * Re16 Pharaoh Creek 82 0/4 768-654 ICE FIELD Pinto Lake Mount E5 Healy Creek 82 0/4 825-608 Re21 Ball Pass Junction 82 0/4 723-652 Mount Sunset Coleman ◊ ◊ Sk5 Hidden Lake 82 N/8 626-029 Saskatchewan Pass E13 Egypt Lake 82 0/4 772-619 Ek13 Elk Lake Summit 82 0/5 951-826 ◊ Sk11 Baker Lake 82 N/8 672-049 Cs Fm10 Mount Cockscomb 82 0/4 923-766 ◊ Sk18 Merlin Meadows 82 N/9 635-093 No 5 ◊ SASKATCHEWAN 11 * Fm19 Mystic Junction 82 0/5 897-834 Sk19 Red Deer Lakes 82 N/9 667-098 River * Fm29 Sawback Lake 82 0/5 868-904 Sf Siffleur 82 N/16 441-356 Mount Gl 9 Glacier Lake 82 N/15 114-528 ◊ Sp6 Mount Rundle 82 0/4 030-647 Amery Alexandra He5 Hector Lake 82 N/9 463-144 Sp16 Rink’s Camp 82 0/4 040-555 Mount Jo9 Larry’s Camp 82 0/5 820-830 * Sp23 Eau Claire 82 J/14 067-505 Wilson * Jo18 Johnston Creek 82 0/5 771-882 * Sp35 Mount Fortune 82 J/14 123-425 ◊ Jo19 Luellen Lake 82 0/5 764-882 Su8 Howard Douglas Lake 82 0/4 880-546 Ta6 Taylor Lake 82 N/8 636-832 SASKATCHEWAN RIVER Jo29 Badger Pass Junction 82 0/5 737-932 N. -

CANADIAN ROCKIES North America | Calgary, Banff, Lake Louise
CANADIAN ROCKIES North America | Calgary, Banff, Lake Louise Canadian Rockies NORTH AMERICA | Calgary, Banff, Lake Louise Season: 2021 Standard 7 DAYS 14 MEALS 17 SITES Roam the Rockies on this Canadian adventure where you’ll explore glacial cliffs, gleaming lakes and churning rapids as you journey deep into this breathtaking area, teeming with nature’s rugged beauty and majesty. CANADIAN ROCKIES North America | Calgary, Banff, Lake Louise Trip Overview 7 DAYS / 6 NIGHTS ACCOMMODATIONS 3 LOCATIONS Fairmont Palliser Calgary, Banff, Lake Louise Fairmont Banff Springs Fairmont Chateau Lake Louise AGES FLIGHT INFORMATION 14 MEALS Minimum Age: 4 Arrive: Calgary Airport (YYC) 6 Breakfasts, 4 Lunch, 4 Dinners Suggested Age: 8+ Return: Calgary Airport (YYC) Adult Exclusive: Ages 18+ CANADIAN ROCKIES North America | Calgary, Banff, Lake Louise DAY 1 CALGARY, ALBERTA Activities Highlights: Dinner Included Arrive in Calgary, Welcome Dinner at the Hotel Fairmont Palliser Arrive in Calgary Land at Calgary Airport (YYC) and be greeted by Adventures by Disney representatives who will help you with your luggage and direct you to your transportation to the hotel. Morning And/Or Afternoon On Your Own in Calgary Spend the morning and/or afternoon—depending on your arrival time—getting to know this cosmopolitan city that still holds on to its ropin’ and ridin’ cowboy roots. Your Adventure Guides will be happy to give recommendations for things to do and see in this gorgeous city in the province of Alberta. Check-In to Hotel Allow your Adventure Guides to check you in while you take time to explore this premiere hotel located in downtown Calgary. -

Pearce Estate Park What We Heard #1
Design Development Plan Phase 2: Pearce Estate Park What We Heard #1: Vision & Programming March–April 2016 _ Report prepared: April 2016 Contents What is Bend in the Bow? 1 Engagement Overview 3 What We Asked 6 What We Heard 7 What We Heard + What We Will Do 12 Next Steps 20 Appendix: Verbatim Comments 21 What is Bend in the Bow? The City of Calgary has begun a long-term project to Phase 1 of this project is completed. It focused on the IBS connect the Inglewood Bird Sanctuary (IBS), Pearce Estate and the Inglewood Wildlands. Please go to Park and the adjoining green spaces along the Bow River— calgary.ca/bendinthebow for a review on what was this project is called Bend in the Bow. discussed and heard at the engagement sessions, and how the preferred design concept evolved. The goal of the project is to explore and address ways to preserve, enhance and celebrate the only urban-centred, Phase 2 of the project is now underway. Phase 2 focuses on federally-recognized bird sanctuary in Canada, while Pearce Estate Park and the adjoining green spaces along the retaining the historic significance of the other lands located Bow River towards the Inglewood Bird Sanctuary. within the new area boundaries. The Design Development Plan (DDP) will integrate the various areas of the two phases Building on Phase 1, Phase 2 will continue with the vision into one cohesive and well-functioning landscape unit. of “a park that tells stories,” with a focus on balancing the core values of nature, culture and education. -

Exploring the Vastness of Banff National Park
Exploring the Vastness of Banff National Park By Claire Walter o borrow on old Ttravelogue cliché, Alberta’s Banff National Park is study in contrast. Its 2,586 square miles comprise both wilderness and civilization. There are high mountains, deep valleys, endless forests and abundant wildlife. Even though much of it feels and looks remote, it is just 70 miles from Calgary – and the Trans-Canada Highway runs right through it. It contains one large town (Banff), one smaller town (Lake Louise Village), two palatial hotels (the Fairmont Banff Springs and Fairmont Chateau Lake Louise) and three significant downhill ski areas (Ski Lake Louise, Sunshine and Norquay). It is a park among parks, with Kootenay National Park just to the south, Yoho National Park to the west (and in another province) and Jasper National Park to the north. It is Canada’s oldest national park and also the one with phenomenal snowshoe opportunities. It’s a great destination for a snowshoe getaway or a multi-activity winter vacation with snowshoeing among the options. There’s skiing (Alpine and Nordic), wildlife viewing, spa- hopping and enjoying the shops, galleries, restaurants and nightspots in Banff or quieter Lake 1 Go FartherTM Model: ARTICA™ BACKCOUNTRY q Two-Piece Articulating Frame q Virtual Pivot Traction Cam q Quick-Cinch™ One-Pull Binding q 80% Recyclable Materials, No PVC’s eastonmountainproducts.com ©2010 easton mountain products Louise Village. As a bonus, winter is low season in Banff, so lodging is a bargain and the shops offer incredible values. Snowshoeing Options The most straightforward snowshoeing is practically from the doorstep of the Chateau Lake Louise.