Factors Influencing Stopover and Movement of Migratory Songbirds Within the Silvio O
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Cavendish Town Plan Select Board Hearing Draft January 2018
Cavendish Town Plan Select Board Hearing Draft January 2018 Town of Cavendish P.O. Box 126 Cavendish, Vermont 05142 (802) 226-7292 Document History • Planning Commission hearing and approval of re-adoption of Town Plan with inclusion of Visual Access Map - February 22, 2012 • Select board review of Planning Commission proposed re-adopted town plan with visual access map - April 9, 2012 • Select board review of town plan draft and approval of SB proposed minor modifications to plan – May 14, 2012 • Planning Commission hearing for re-adoption of Town Plan with Select board proposed minor modifications – June 6, 2012 • Planning Commission Approval of Re-adoption of Town Plan with minor modifications – June 6, 2012 • 1st Select board hearing for re-adoption of town plan with minor modifications – June 11, 2012 • 2nd Select board Hearing for re-adoption of Town Plan with minor modifications – August 20, 2012 • Cavendish Town Plan Re-adopted by Australian ballot at Special Town Meeting – August 28, 2012 • Confirmation of Planning Process and Act 200 Approval by the Southern Windsor County Regional Planning Commission – November 27, 2012 • Planning Commission is prepared updates in 2016-2017 This report was developed in 2016 and 2017 for the Town of Cavendish with assistance from the Southern Windsor County Regional Planning Commission, Ascutney, VT. Financial support for undertaking this revision was provided, in part, by a Municipal Planning Grant from the Vermont Agency of Commerce and Community Development. ii Table of Contents Chapter 1: Introduction ............................................................................................................... 1 1.1 Purpose ............................................................................................................................. 1 1.2 Planning Process Summary................................................................................................... 3 1.2 Community and Demographic Trends ............................................................................ -

Okemo State Forest - Healdville Trail Forest - Healdville Okemo State B
OKEMO STATE FOREST - HEALDVILLE TRAIL North 3000 OKEMO MOUNTAIN RESORT SKI LEASEHOLD AREA OKEMO MOUNTAIN ROAD (paved) 2500 2000 Coleman Brook HEALDVILLE TRAIL 1500 to Ludlow - 5 miles STATION RD railroad tracks HEALDVILLE RD HEALDVILLE VERMONT UTTERMILK F 103 B AL LS RD to Rutland - 16 miles Buttermilk Falls 0 500 1000 2000 3000 feet 1500 LEGEND Foot trail Vista Town highway State highway Lookout tower FORESTS, PARKS & RECREATION State forest highway (not maintained Parking area (not maintained in winter) VERMONT in winter) Gate, barricade Stream AGENCY OF NATURAL RESOURCES Ski chairlift Ski area leasehold boundary 02/2013-ephelps Healdville Trail - Okemo State Forest the property in 1935. Construction projects by the CCC The Healdville Trail climbs from the base to the include the fire tower, a ranger’s cabin and an automobile summit of Okemo Mountain in Ludlow and Mount Holly. access road. The majority of Okemo Mountain Resort’s Highlights of this trail include the former fire lookout ski terrain is located within a leased portion of Okemo tower on the summit and a vista along the trail with State Forest. Okemo State Forest is managed for Okemo views to the north and west. Crews from the Vermont multiple uses under a long-term management plan; these Youth Conservation Corps constructed the trail under the uses include forest products, recreation and wildlife direction of the Vermont Department of Forests, Parks habitat. Okemo State Forest provides an important State Forest and Recreation during the summers of 1991-1993. wildlife corridor between Green Mountain National Forest lands to the south and Coolidge State Forest to the Trail Facts north. -

H.880: an Act Relating to Abenaki Place Names on State Park Signs
H.880: An act relating to Abenaki place names on State park signs Representative Brian Cina September 8, 2020 H.880: An act relating to Abenaki place names on State park signs By adding Abenaki place names alongside colonial place names on State park signs, the State of Vermont intends to: - Recognize that the State of Vermont exists on territory originally and currently inhabited by Abenaki people. - Increase visibility and awareness of the Abenaki people and culture. - Preserve and promote the Abenaki language. - Honor the history, significance, and spirit of places. Examples of Abenaki place names: Abenaki Colonial meaning of Abenaki name Odzhihozo Rock Dunder “he who created himself” Bitawbakw Lake Champlain "in between water" Winoski Winooski "onion land" Mazipskoik Missisquoi "place of flint" Wnekikwisibo Otter Creek "otter river/creek" Kwenitekw Connecticut River "long flow (river)" Kchi Pontekw Bellows Falls "great falls" Tawipodiwajo, also Moziozagan Camel's Hump "place to sit in mountain" Mozdebiwajo Mount Mansfield "moosehead mountain" Gwenaden Jay Peak "long mountain" Kaskaden Mount Ascutney "wide mountain" Mategwasaden Mount Philo "rabbit mountain" Holschuh, Rich (2019, October 17). Mountain Names: Remembering Their Aboriginal Origins. Retrieved February 25, 2020 from: https://www.greenmountainclub.org/mountain-names-remembering-their-aboriginal-origins/ Possible Witnesses for H.880 Vermont Commission on Native American Affairs Elnu Abenaki Tribe Nulhegan Band of the Coosuk Abenaki Nation Koasek Abenaki of the Koas Missisquoi, St. Francis-Sokoki Band Vermont Historical Society Commissioner of Forest, Parks, and Recreation Questions? nadodmowôganal?. -

Preliminary Bedrock Geologic Map of the Chittenden Quadrangle Rutland County, Vermont
U.S. DEPARTMENT OF THE INTERIOR U. S. GEOLOGICAL SURVEY Preliminary Bedrock Geologic Map of the Chittenden Quadrangle Rutland County, Vermont By Nicholas M. Ratcliffe U.S. Geological Survey Open File Report 97-703 1997 This report is preliminary and has not been reviewed for conformity with U.S. Geological Survey editorial standards nor with the North American Stratigraphic Code. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. INTRODUCTION The bedrock of the Chittenden quadrangle consists of Middle Proterozoic through Cambrian metasedimentary and metaigneous rocks that lie along the western flank of the Green Mountain massif in south-central Vermont. The Green Mountain massif is cored by highly deformed and extensively retrograded schists and schistose gneisses derived from once high-grade gneisses that were intruded and originally metamorphosed during the Grenville orogeny prior to about 950 Ma. Upper Proterozoic through Cambrian metaclastic rocks, referred to as the western cover sequence, form a cover sequence that was deposited unconformably on the older schists and gneisses. The upper part of the cover sequence consists of marine-shelf carbonate rocks of the Vermont Valley sequence that developed on the stable Atlantic-type continental margin prior to the deformation in the Taconian orogeny during the Middle to Upper Ordovician. In the Taconian orogeny, rocks of this area were thrust faulted and metamorphosed to biotite grade. The basement rocks were altered by low-grade remetamorphism and hydration to such an extent that the Grenvillian mineral assemblages, schistosity, and gneissosity are presently barely recognizable. -

Summer 2021 Long Trail News
NEWS Quarterly of the Green Mountain Club SUMMER 2021 555 New End-to-Enders The Most Ever Recorded [FROM AGE 7 TO 80] CONTENTS Summer 2021, Volume 81, No. 2 The mission of the Green Mountain Club is to make the Vermont mountains play a larger part in the life of the people by protecting and maintaining the Long Trail System and fostering, through education, the stewardship of Vermont’s hiking trails and mountains. Quarterly of the Green Mountain Club Michael DeBonis, Executive Director Chloe Miller, Communications Manager & Long Trail News Editor Richard Andrews, Volunteer Copy Editor Sylvie Vidrine, Graphic Designer Green Mountain Club 4711 Waterbury-Stowe Road Waterbury Center, Vermont 05677 Phone: (802) 244-7037 Fax: (802) 244-5867 E-mail: [email protected] Website: greenmountainclub.org The Long Trail News is published by The Green Mountain Club, Inc., a nonprofit organization founded in 1910. In a 1971 Joint Resolution, the Vermont Legislature Julia LanzDuret-Hernandez on Mount Mansfield. designated the Green Mountain Club the “founder, sponsor, defender and protector of the Long Trail System...” FEATURES Contributions of manuscripts, photos, illustrations, and news are welcome from ❯ The Mountains That Made Me members and nonmembers. 5 by Julia LanzDuret-Hernandez The opinions expressed byLTN contributors are not necessarily those of GMC. 6 ❯ Congratulations to the 2020 End-to-Enders The Long Trail News (USPS 318-840) is published quarterly by The Green Mountain and Side-to-Siders Club, Inc., 4711 Waterbury-Stowe Road, ❯ Waterbury Center, VT 05677. Periodicals 8 A Day in the Life: Long Trail Thru-Hiker postage paid at Waterbury Center, VT, and additional offices. -

This Is the Bennington Museum Library's “History-Biography” File, with Information of Regional Relevance Accumulated O
This is the Bennington Museum library’s “history-biography” file, with information of regional relevance accumulated over many years. Descriptions here attempt to summarize the contents of each file. The library also has two other large files of family research and of sixty years of genealogical correspondence, which are not yet available online. Abenaki Nation. Missisquoi fishing rights in Vermont; State of Vermont vs Harold St. Francis, et al.; “The Abenakis: Aborigines of Vermont, Part II” (top page only) by Stephen Laurent. Abercrombie Expedition. General James Abercrombie; French and Indian Wars; Fort Ticonderoga. “The Abercrombie Expedition” by Russell Bellico Adirondack Life, Vol. XIV, No. 4, July-August 1983. Academies. Reproduction of subscription form Bennington, Vermont (April 5, 1773) to build a school house by September 20, and committee to supervise the construction north of the Meeting House to consist of three men including Ebenezer Wood and Elijah Dewey; “An 18th century schoolhouse,” by Ruth Levin, Bennington Banner (May 27, 1981), cites and reproduces April 5, 1773 school house subscription form; “Bennington's early academies,” by Joseph Parks, Bennington Banner (May 10, 1975); “Just Pokin' Around,” by Agnes Rockwood, Bennington Banner (June 15, 1973), re: history of Bennington Graded School Building (1914), between Park and School Streets; “Yankee article features Ben Thompson, MAU designer,” Bennington Banner (December 13, 1976); “The fall term of Bennington Academy will commence (duration of term and tuition) . ,” Vermont Gazette, (September 16, 1834); “Miss Boll of Massachusetts, has opened a boarding school . ,” Bennington Newsletter (August 5, 1812; “Mrs. Holland has opened a boarding school in Bennington . .,” Green Mountain Farmer (January 11, 1811); “Mr. -
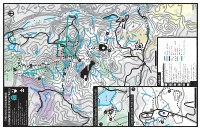
THE C OOLIDGE R ANGE SUMMER RECREATION TR AILS Legend Giff
VERMONT THE COOLIDGE RANGE Long Trail North 100 Tucker Johnson 2000 Thundering North SUMMER RECREATION TRAILS Brook Rd Willard Gap Giord Woods Kent Pond Coolidge State Forest State Park Giord Woods State Park 3 Coolidge State Park Deer Leap Mtn Plymsbury Wildlife Management Area 2782’ Rd River Green Mountain National Forest Deer Leap Old Maine Jct. Appalachain Trail Corridor Overlook Rutland City Forest 2000 k West Hill Rd o o Forest Legacy Public Access Easments r B t n e 4 K 4 Killington Rd Pico Pond VERMONT 3000 100 3 Wheelerville Rd 4 3800 Churchill Scott Pico Camp/ spring Pico Peak 3957’ Little Pico Gre o k 3110’ at R oaring Bro Ottauquechee River Rams Head Mtn 3618’ East Roaring Brook Rd Brewers Corners Shagback Mtn Brewers Brook 7 Snowdon Peak 2688’ 3592’ 1800 Skye Peak 2000 3816’ 1600 2200 Cooper Lodge Ed dy 2400 Bro ok 2600 4 Killington Peak ok 2800 s ro 4235’ Fall B 3000 3 Bear Mtn Wheelerville Rd Ottauquechee 3262’ River Notch Rd Mendon Peak 3800 3840’ 3600 Little Killington Peak 3939’ 3200 Ma 3000 dden Brook Reservoir Brook Giord Woods State Park Trails VERMONT North VERMONT Shrewsbury Peak 100 100 3710’ Smith Peak ok Robinson Hill ro 3205’ B t n 2747’ e Shrewsbury Peak rg a S 6 Kent Pond 3200 9 Gov. Clement 3000 1 2800 5 2600 Woodard Jockey Hill Reservoir 2400 2640’ Russell Stone Hut Hill CCC Road 1800 Ingalls Hill Russell Hill 1600 2654’ 2545’ 2000 Tinker Brook 8 Black Pond ko Thundering T ro Brook Rd i n B 2200 k e r 1000 500 0 1000 2000 4 feet Tin Shanty Rd Shanty Tin Black River North Coolidge State Park Trails Upper Cold River Rd Burnt Mtn 2803’ VERMONT 1200 100A 2000 Cold River Rd 2 Northam Rd Round Top Mtn Rd 1400 VERMONT North Shrewsbury Old Plymouth Rd 100 k to Coolidge State Park oo Br ing via Rt 100A 3 mi. -

The Mount Ascutney Parkway
The Mount Ascutney Parkway The Mount Ascutney Parkway is a 3.7-mile road built by hand and machine by the South Peak Civilian Conservation Corps from 1933 to 1937. The CCC established a camp on property acquired for conservation and recreation projects in 1933. During the six year North time the camp was active, crews worked on surveying and constructing the parkway, building hiking trails, a campground, a ski jump, picnic sites and forestry projects. Road construction included drilling and blasting much of the route using steam drills, hand tools and some trucks and equipment. The road was paved in the 1960’s concurrent with campground expansion. Today, over 15,000 people enjoy the parkway in the summer Windsor months to take in long-range views, hike the summit, camp at the base and participate vista in foot and bicycle races and auto time trials. Hours and scheduled road closures are posted on the park’s webpage. Mount Ascutney rises 2,500 ft. from the Connecticut River Valley to an elevation of 3,144ft. above sea level, forming a dominating topographical feature visible in both southeastern Vermont and southwestern New Hampshire. Grades average 10% on the parkway with some sections as steep at 19%. Drive Safely Drive all the way up and down in low gear. Using the lowest gear keeps stress on your vehicle’s transmission to a minimum and helps control your speed on the way down. Low gear can be denoted by an “L,” “L1” or “1” or “M1.” Ascending in low gear places less load on your engine and allows it to run cooler. -

Vermont's Guide to Fluoride Levels in Public Water Systems
Vermont’s Guide to Fluoride Levels in Public Water Systems May 2015 Oral Health healthvermont.gov May 2015 The Vermont Guide to Fluoride Levels in Public Water Systems is prepared by the Vermont Department of Health, and updated every three years. This 2015 edition replaces previous versions. For more information or assistance, please contact the Office of Oral Health, Vermont Department of Health at 802-651-1635 or 866-331-5622. Recommended Fluoride Level The current recommended level for fluoride in public water systems is 0.70 parts per million (ppm). For Primary Health Care Providers Primary health care providers (pediatricians, family physicians, physician assistants, nurse practitioners, etc.) and dentists make decisions daily as to the need for prescribing fluoride supplements for children. The information in this guide will enable providers to determine at a glance, the level of fluoride the family is already receiving from their community water system prior to prescribing any supplementation. If the family is served by a private well or spring system, their water must be tested prior to prescribing a dietary fluoride supplement. The Vermont Department of Health (VDH) encourages primary care providers to identify children at risk for Early Childhood Caries (ECC) and to help ensure that those children have access to early preventive care. A key preventive measure for ECC is the proper use of fluoride, both systemically and topically. Fluoride makes a tooth more resistant to decay, inhibits the bacteria’s ability to create acid, and promotes remineralization of white spot lesions (early decay). These effects on teeth are both pre-eruptive and post- eruptive. -

Become a Vermont State Parks Junior Ranger Help a Ranger What Do You Need to Do?
Become a Vermont State Parks Junior Ranger Help a Ranger What do you need to do? □ Visit Vermont State Parks. Directions: Read these questions and circle the best answer listed below. □ Care about taking care of Vermont State Parks. 1) You notice someone carving initials in the wall of a lean-to. Is this: A) A special way to commemorate a camping trip. □ Complete the required pages in this booklet: B) Graffiti that will damage the lean-to and make it look yucky. □ If you are between ages 4 and 8, complete all the pages with a salamander picture at 2) You see a beautiful flower on the side of a trail in a state park. So you: the top and you will become a Salamander A) Pick it to take it with you and share with others. Level Junior Ranger. B) Take a picture of it to share with others. If you are age 9 years or older, complete C) Remember where it is so you can take people to that spot and □ share it. all the pages with a Beaver picture at the top and you will become a Beaver Level D) Stomp on it with your foot. Junior Ranger. 3) Before you go on a hike, you need to pack: (select all that apply) A) Healthy snacks □ Bring your completed booklet to park staff in any Vermont State Park. B) Water C) A bowling ball □ Take the Junior Ranger pledge. D) Extra layers of clothing _________________________________________________ Take Action: help your favorite park by doing a service project. -

Green Mountains, Vermont Postcard Folder, 1940S
Green Mountains, Vermont Fairchild Aerial Surveys, Inc., Postcard Folder – 1940’s This article, which begins on the next page, is presented on the Stone Quarries and Beyond web site. http://quarriesandbeyond.org/ Peggy B. Perazzo Email: [email protected] September 2012 Bird’s Eye View, Battle Monument, Bennington, VT. Looking towards Wilmington from Fire Tower on Mount Olga, Molly Stark Trail, Brattleboro Molly Stark Trail Looking Toward Bennington, VT. Mt. Equinox, Manchester, VT. Green Mts. View from “The Brow” of Green Mts. and Pownal Valley, Pownal, Vt. Emerald Lake in the Green Mountains, Vermont Airplane View of Lake Bomoseen in the Green Mts., Vermont Chittenden Dam in the Green Mountains, Vermont Bird’s-Eye View, Lake Dunmore, Green Mts. Hortonia, Echo and Bebee Lakes in the Green Mts. of Vermont Camel’s Hump, Green Mountains, Vermont Mt. Mansfield and Lamoille River, Green Mts. of Vermont Mt. Mansfield, VT. Altitude 4364 Feet. Highest peak in the Green Mountains. Where the road winds between giant boulders in Smugglers Notch, Green Mts., VT. Covered Bridge, West Brattleboro, Ct., in the Green Mountains Bird’s-Eye View of Montpelier, VT. Showing State Capitol. Looking toward Northfield, from Allis State Park, Green Mts., Vt. Mount Ascutney and Connecticut River Below is a transcription of the introduction to this postcard folder with views of Vermont, “The Green Mountain State”: The Green Mountain State – Historical Places of Interest Brattleboro: Location of Fort Dummer, 1724, first permanent settlement in Vermont: former home of Rudyard Kipling. Montpelier: State Capitol. State House contains man interesting relics. Birthplace of Admiral Dewey. -

Green Mountains, Vermont
Green Mountains, Vermont Fairchild Aerial Surveys, Inc., Postcard Folder – 1940’s This article, which begins on the next page, is presented on the Stone Quarries and Beyond web site. http://quarriesandbeyond.org/ Peggy B. Perazzo Email: [email protected] September 2012 Bird’s Eye View, Battle Monument, Bennington, VT. Looking towards Wilmington from Fire Tower on Mount Olga, Molly Stark Trail, Brattleboro Molly Stark Trail Looking Toward Bennington, VT. Mt. Equinox, Manchester, VT. Green Mts. View from “The Brow” of Green Mts. and Pownal Valley, Pownal, Vt. Emerald Lake in the Green Mountains, Vermont Airplane View of Lake Bomoseen in the Green Mts., Vermont Chittenden Dam in the Green Mountains, Vermont Bird’s-Eye View, Lake Dunmore, Green Mts. Hortonia, Echo and Bebee Lakes in the Green Mts. of Vermont Camel’s Hump, Green Mountains, Vermont Mt. Mansfield and Lamoille River, Green Mts. of Vermont Mt. Mansfield, VT. Altitude 4364 Feet. Highest peak in the Green Mountains. Where the road winds between giant boulders in Smugglers Notch, Green Mts., VT. Covered Bridge, West Brattleboro, Ct., in the Green Mountains Bird’s-Eye View of Montpelier, VT. Showing State Capitol. Looking toward Northfield, from Allis State Park, Green Mts., Vt. Mount Ascutney and Connecticut River Below is a transcription of the introduction to this postcard folder with views of Vermont, “The Green Mountain State”: The Green Mountain State – Historical Places of Interest Brattleboro: Location of Fort Dummer, 1724, first permanent settlement in Vermont: former home of Rudyard Kipling. Montpelier: State Capitol. State House contains man interesting relics. Birthplace of Admiral Dewey.