Pinyon-Juniper Woodland Classification and Description Research Natural Areas of Southeastern Idaho
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

U.S. Geological Survey Radiometric Ages—Compilation "C" Part Four: Idaho, Oregon, and Washington R.H
U.S. Geological Survey radiometric ages—compilation "C" Part four: Idaho, Oregon, and Washington R.H. Marvin, H.H. Mehnert, C.W. Naeser, and R.E. Zartman Isochron/West, Bulletin of Isotopic Geochronology, v. 53, pp. 3-13 Downloaded from: https://geoinfo.nmt.edu/publications/periodicals/isochronwest/home.cfml?Issue=53 Isochron/West was published at irregular intervals from 1971 to 1996. The journal was patterned after the journal Radiocarbon and covered isotopic age-dating (except carbon-14) on rocks and minerals from the Western Hemisphere. Initially, the geographic scope of papers was restricted to the western half of the United States, but was later expanded. The journal was sponsored and staffed by the New Mexico Bureau of Mines (now Geology) & Mineral Resources and the Nevada Bureau of Mines & Geology. All back-issue papers are available for free: https://geoinfo.nmt.edu/publications/periodicals/isochronwest This page is intentionally left blank to maintain order of facing pages. U.S. GEOLOGICAL SURVEY RADIOMETRIC AGES-COMPILATION "C" Part four: Idaho, Oregon, and Washington RICHARD F. MARVIN HARALD H. MEHNERT U.S. Geological Survey, Federal Center, Denver, CO 80225 CHARLES W. NAESER ROBERT E. ZARTMAN This is part four of the third compilation of a planned series of age determinations by the U.S. Geological Survey appearing in Isochron/West. It contains both unpublished and published dates. However,the latter ages lacked either a specific sample location, petrologic information, or ana lytical data in their published form. Such specifics are often needed for evaluation and utilization of an isotopic age. That, we believe, is the value of this compilation—it sup plies such information for most of the listed samples. -

Payette National Forest
Appendix 2 Proposed Forest Plan Amendments Sawtooth National Forest Land and Resource Management Plan Chapter III Sawtooth WCS Appendix 2 Chapter III. Management Direction Table of Contents Management Direction......................................................................................................... III-1 Forest-Wide Management Direction ................................................................................ III-1 Threatened, Endangered, Proposed, and Candidate Species ....................................... III-1 Air Quality and Smoke Management .......................................................................... III-4 Wildlife Resources ....................................................................................................... III-5 Vegetation .................................................................................................................... III-9 Non-native Plants ....................................................................................................... III-13 Fire Management ....................................................................................................... III-14 Timberland Resources ............................................................................................... III-16 Rangeland Resources ................................................................................................. III-17 Minerals and Geology Resources .............................................................................. III-18 Lands and Special -

CAI) As an Aid to Structural Interpretation in the Black Pine Mountains, Idaho
Conodont Color Alteration (CAI) as an Aid to Structural Interpretation in the Black Pine Mountains, Idaho By J. Fred Smith, Jr. and Bruce R. Wardlaw Open-File Report 2012–1100 U.S. Department of the Interior U.S. Geological Survey U.S. Department of the Interior KEN SALAZAR, Secretary U.S. Geological Survey Marcia K. McNutt, Director U.S. Geological Survey, Reston, Virginia: 2012 For product and ordering information: World Wide Web: http://www.usgs.gov/pubprod Telephone: 1–888–ASK–USGS For more information on the USGS—the Federal source for science about the Earth, its natural and living resources, natural hazards, and the environment: World Wide Web: http://www.usgs.gov Telephone: 1–888–ASK–USGS Suggested citation: Smith, J.F., Jr., and Wardlaw, B.R., 2012, Conodont color alteration (CAI) as an aid to structural interpretation in the Black Pine Mountains, Idaho: U.S. Geological Survey Open-File Report 2012–1100, 5 p., available only at http://pubs.usgs.gov/of/2012/1100. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government. Although this report is in the public domain, permission must be secured from the individual copyright owners to reproduce any copyrighted material contained within this report. ii Contents Abstract ......................................................................................................................................................................... 1 Introduction ................................................................................................................................................................... -

Baseline and Stewardship Monitoring on Sawtooth National Forest Research Natural Areas
Baseline and stewardship monitoring on Sawtooth National Forest Research Natural Areas Steven K. Rust and Jennifer J. Miller April 2003 Idaho Conservation Data Center Department of Fish and Game 600 South Walnut, P.O. Box 25 Boise, Idaho 83707 Steven M. Huffaker, Director Prepared for: USDA Forest Service Sawtooth National Forest ii Table of Contents Introduction ............................................... 1 Study Area ............................................... 1 Methods ................................................. 4 Results .................................................. 5 Recommendations and Conclusions .......................... 12 Literature Cited ........................................... 14 List of Figures ............................................ 16 List of Tables ............................................ 26 Appendix A .............................................. 35 Appendix B .............................................. 36 Appendix C .............................................. 61 iii iv Introduction Research natural areas are part of a national network of ecological areas designated in perpetuity for research and education and to maintain biological diversity on National Forest System lands. Seven research natural areas occur on Sawtooth National Forest: Basin Gulch, Mount Harrison, Pole Canyon, Pole Creek Exclosure, Redfish Lake Moraine, Sawtooth Valley Peatlands, and Trapper Creek (Figure 1). These natural areas were established in the late 1980s and mid 1990s to provide representation of a diverse -

IBT Hikers Guide PDF 6X9 Paper Version
IDAHO BOUNDARY TRAIL HIKERS GUIDE 2020 By MIKE O’BRIEN & Jim “Yeti” Fulmis Copyright © 2020 www.fultek.com/ibt Please note that any discrepancies such as US Post office hour changes, prices, businesses closed or burned down is way beyond our control. If you discover changes, please send us information via email ([email protected]). This will help us to keep this guide up to date as best as possible. We did the trail in 2016 and updated everything, including all tracks and waypoints, which can be found at www.fultek.com/ibt. Trail closures, fires, or storms that blow down a lot of trees can and will happen. We discovered that ourselves in 2016. If you can, track anything you find different or just describe the differences and send those to the above email address. Tracks should be in GPX form. Also, there will be a text file named IBT_Hikers_Guide_Updates.txt online at www.fultek.com/ibt for any changes or additions we find out about after the book is published. Cover Photo: On the way to Owyhee State Park. Copyright © 2020 Mike O’Brien & Jim “Yeti” Fulmis Edited by: Melanie Simmerman All rights reserved. ISBN-13: 978-1793257536 Why another long trail? When we looked at a map of Idaho we saw a wide variety of terrain, desert, forest, mountains; lots of mountains. Not only that, but the border was mostly public land where we could choose where to hike and camp. Could we hike all the way around? It would be tough. First, we had to figure out a route, one that had reliable water in the desert (no trail angels in the Owyhee Desert!), and a route that touched the special places, like the Tetons, the Bitterroot and Selkirk Mountains, the cedar rainforests in the north. -

Paleozoic Rocks in the Black Pine Mountains, Cassia County, Idaho
Paleozoic Rocks in the Black Pine Mountains, Cassia County, Idaho GEOLOGICAL SURVEY BULLETIN 1536 Paleozoic Rocks in the Black Pine Mountains, Cassia County, Idaho By]. FRED SMITH, JR. GEOLOGICAL SURVEY BULLETIN 1536 Descriptions of eleven rock units which range in age from Devonian to Permian, the thickest parts being Early Pennsylvanian to· Early Permian UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON 1983 UNITED STATES DEPARTMENT OF THE INTERIOR JAMES G. WATT, Secretary GEOLOGICAL SURVEY Dallas L. Peck, Director Library of Congress Cataloging in Publication Data Smith, J. Fred (Joe Fred), 1911-1982 Paleowic rocks in the Black Pine Mountains, Cassia County, Idaho. (Geological Survey Bulletin 1536) Bibliography: 36 p. Supt. of Docs. No.: I 19.3: 1. Geology, Stratigraphic-Paleozoic. 2. Geology-Idaho-Black Pine Mountains. I. Title. II. Series. QE75.B9 no. 1536 557.3s [551.7'2'0979639] 81-607193 [QE.654] AACR2 For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C. 20402 CONTENTS Page Abstract . 1 Introduction . 2 Stratigraphy . 7 Rocks south of West Dry Canyon fault . 8 Lower plate . 8 Devonian System . 8 Jefferson Formation . 8 Mississippian and Pennsylvanian Systems . 9 Manning Canyon Shale . 9 Middle plate . 11 Pennsylvanian System . 11 Oquirrh Formation, limestone member . 11 Lower part . 11 Middle part . 13 Upper part . 15 Oquirrh Formation, limestone, sandstone, and quartzite mem- ber . 17 Oquirrh Formation, limestone and dolomite member . 18 Upper plate . 21 Pennsylvanian and Permian Systems . 21 Oquirrh Formation sandstone and siltstone member . 21 Rocks north of West Dry Canyon fault . 23 Plate I . 23 Pennsylvanian and Permian Systems . -
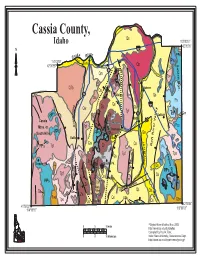
Cassia County, Qs Idaho 113O0010 O Qw 42 3505 86 Qw Ke Iver Ppsp Sna R O Pps 114 0350 Qa Burley O Qb Sublett Rge
Cassia County, Qs Idaho 113o0010 o Qw 42 3505 86 Qw ke iver PPsP Sna R o PPs 114 0350 Qa Burley o Qb Sublett Rge. 42 3055 Declo Qs 77 PPsP Qm PPsP Tov Zs Wm QTb Qa PPs QTb Tpf Idahome PPsP Qs Qs PPsP 27 Albion Tps Qs QTb Ps Qs 84 k Zs e Qw Qs e r Tpf 81 TsR Qw C Zs Sublett P Tpf Wm PPs e Malta Qs s Tov Cassia o Wm o Zs Wm Pzu G Tpf Mtns. or Mt. Harrison South Hills PPsP PPsP PPsP Qa Oakley PzZm Elba Ps Ps Qs Qs r e Pzu Tpf Zs Raft River Valley Ps PPsP Ps Blackpine Rge. v Zs i R Tps Tpf Qs Tpf Wm t PzZm f PPs Tpf Zs Tpf a Trapper Pk. Cache Pk. Tms R Qa Tpf Toi Toi PPsP Tpf Zs Tpf Toi Qs Qs Almo Tpf Ms PPsP Tpf Albion Mtns. Wm PPs Zs Zs Toi PPs PPs Almo Qs Qa PzZm Pluton Raft RiverTpf Narrows. Qa Tpf Zs City of Tms Qs Tps Tpf PPsP Qbo Qs Rocks Tpf Qs o o 42 0004 41 5932 o 114o1655 113 0005 Digital Atlas of Idaho, Nov. 2002 0 5 10 miles http://imnh.isu.edu/digitalatlas Compiled by Paul K. Link, 0 8 16 kilometers Idaho State University, Geosciences Dept. http://www.isu.edu/departments/geology/ Cassia County Cassia County, on the south side of the Snake River forms much of Idahos southern boundary with Utah and Nevada on the west. It contains a diverse assemblage of rocks, including the oldest rocks in Idaho, the metamorphic Green Creek gneiss in the Albion Mountains core complex. -

Albion Raft River and Grouse Creek Field Trip Guide
1 THE ALBION-RAFT RIVER-GROUSE CREEK METAMORPHIC CORE COMPLEX: GEOLOGIC SETTING AND FIELD TRIP GUIDE Elizabeth Miller, Ariel Strickland and Alex Konstantinou, Dept. Geological and Environmental Sciences, Stanford University (Revised from: Elizabeth Miller and Ariel Strickland, 2007 Stanford Field Trip to ARG. Caution: Not necessarily accurate or up-to-date!!!!) Introduction The Albion-Raft River-Grouse Creek metamorphic core complex (ARG) is part of a chain of Cordilleran metamorphic core complexes that lie to the west of the Sevier fold- and-thrust belt (Fig. 1). The ARG is made up of three mountain ranges for which it is named (Fig. 2). The Albion Mountains and Grouse Creek Mountains are characteristic of mountain ranges in the Basin and Range Province; they have a generally north-south orientation and rise abruptly from the surrounding relatively flat topography. The Raft River Mountains have an enigmatic east-west trend and project eastward from the northern end of the Grouse Creek Mountains (Fig. 2). Late Archean crystalline basement is exposed in all three mountain ranges indicating a broad dome or structural culmination beneath this region, where probably 10-15 km minimum of relative vertical uplift (compared to surrounding regions) has occurred. In addition, significant thinning of the overlying upper part of the crust characterizes the geology of this core complex. Eocene and Oligocene granitic plutons are exposed in these ranges and based on geologic relations and cross-sections, most likely underlie much of the ARG. The ARG is unique in the Basin and Range because it is bound on both of its sides (east and west) by normal fault systems and their associated basins. -

Geologic Map of the Kelton Pass Quadrangle, Box Elder County, Utah, and Cassia County, Idaho
GEOLOGIC MAP OF THE KELTON PASS QUADRANGLE, BOX ELDER COUNTY, UTAH, AND CASSIA COUNTY, IDAHO by Michael L. Wells Department of Geoscience, University of Nevada, Las Vegas 4505 S. Maryland Parkway, Las Vegas, NV 89154-4010 Cover Photo: Crystal Peak and the north side of upper Ten Mile Canyon, looking to the northeast. The Raft River detachment is marked by black ledge of resistant fault rocks, overlying the prominent gently dipping white Elba Quartzite. Photo by Michael L. Wells. ISBN 1-55791-807-4 MISCELLANEOUS PUBLICATION 09-3 UTAH GEOLOGICAL SURVEY a division of Utah Department of Natural Resources 2009 STATE OF UTAH Jon Huntsman, Jr., Governor DEPARTMENT OF NATURAL RESOURCES Michael Styler, Executive Director UTAH GEOLOGICAL SURVEY Richard G. Allis, Director PUBLICATIONS contact Natural Resources Map & Bookstore 1594 W. North Temple Salt Lake City, Utah 84116 telephone: 801-537-3320 toll-free: 1-888-UTAH MAP Web site: mapstore.utah.gov email: [email protected] UTAH GEOLOGICAL SURVEY contact 1594 W. North Temple, Suite 3110 Salt Lake City, Utah 84116 telephone: 801-537-3300 fax: 801-537-3400 Web site: geology.utah.gov The Miscellaneous Publication series provides non-UGS authors with a high-quality format for documents concerning Utah geology. Although review comments have been incorporated, this publication does not necessarily conform to UGS technical, policy, or editorial standards. The Utah Department of Natural Resources, Utah Geological Survey, makes no warranty, expressed or implied, regarding the suitability of this product for a particular use. The Utah Department of Natural Resources, Utah Geological Survey, shall not be liable under any circumstances for any direct, indirect, special, incidental, or consequential damages with respect to claims by users of this product. -

Geology and Mineralization of the Southeastern Part of the Black Pine Mountains, Cassia County, Idaho
Utah State University DigitalCommons@USU All Graduate Theses and Dissertations Graduate Studies 5-1975 Geology and Mineralization of the Southeastern Part of the Black Pine Mountains, Cassia County, Idaho Don E. French Utah State University Follow this and additional works at: https://digitalcommons.usu.edu/etd Part of the Geology Commons Recommended Citation French, Don E., "Geology and Mineralization of the Southeastern Part of the Black Pine Mountains, Cassia County, Idaho" (1975). All Graduate Theses and Dissertations. 1903. https://digitalcommons.usu.edu/etd/1903 This Thesis is brought to you for free and open access by the Graduate Studies at DigitalCommons@USU. It has been accepted for inclusion in All Graduate Theses and Dissertations by an authorized administrator of DigitalCommons@USU. For more information, please contact [email protected]. GEOLOGY AND MINERALIZATION OF THE SOUTHEASTERN PART OF THE BLACK PINE MOUNTAINS, CASSIA COUNTY, IDAHO by Don E. French A thesis submitted in partial fulfillment of requirements for the degree of MASTER OF SCIENCE in Geology Approved: Major Professor Committee Member Committee Member Dean of Graduate Studies UTAH STATE UNIVERSITY Logan, utah 1975 ii ACKNOWLEDGMENTS The author is grateful to Dr. Donald R. Olsen, Dr. Clyde T. Hardy, and Dr. Richard R. Alexander for their assistance in preparation of this report. Helpful comments were also given by Mr. Richard Harris and Mr. Perry West of Newmont Exploration Ltd. and by Mr. Louis Cramer of Salt Lake City, Utah. An important contribution was made by Patricia S. French who provided en couragement and financial and technical assistance. Don E. French iii TABLE OF CONTENTS Page ACKNOWLEDGMENTS · . -

Black Pine Management Area 19
Chapter III Black Pine Management Area 19 6.1 ¨¦§84 6.1 on Cany on rock ny Sand a s C ne Jo 6.1 Juniper " 6.1 761 3.2 5 86 2.2 Kelsaw Canyon Black " Pine 6.1 n Black Pine IRA o y n a C e n i P k 7 c 8 a l 5 B revell Canyon 771 St A H O " 2 I D Strevell 8 0 U T A H 02468Miles Legend Management Prescription Categories ¯ 2.2 Research Natural Area 3.2 Active Restoration and Maintenance of Aquatic, Terrestrial, and Hydrologic Resources 6.1 Restoration and Maintenance Emphasis within Shrubland and Grassland Landscapes The Forest Service uses the most current and complete Non-Forest System Lands data available. GIS data and product accuracy may vary. Inventoried Roadless Areas (IRAs) Using GIS products for purposes other than those intended may yield inaccurate or misleading results. Map produced by: B.Geesey, Sawtooth NF, 09/2009 Management Area 19: Black Pine Location Map III - 335 Chapter III Black Pine Management Area 19 Management Area 19 Black Pine MANAGEMENT AREA DESCRIPTION Management Prescriptions - Management Area 19 has the following management prescriptions (see map on preceding page for distribution of prescriptions). Percent of Management Prescription Category (MPC) Mgt. Area 2.2 – Research Natural Areas Trace 3.2 – Active Restoration and Maintenance of Aquatic, Terrestrial & Hydrologic Resources 59 6.1 – Restoration and Maintenance Emphasis within Shrubland & Grassland Landscapes 41 General Location and Description - Management Area 19 is comprised of Forest Service lands in the Black Pine Mountain Range (see map, preceding page), which lies at the eastern end of Cassia County, Idaho. -

Idaho State Plan 2000
Idaho Partners in Flight Idaho Bird Conservation Plan Version 1.0 January 2000 Address Questions to: Sharon Ritter Idaho Partners in Flight Coordinator 142 West Hills Way Hamilton, MT 59840 406-375-9150 [email protected] EXECUTIVE SUMMARY JUSTIFICATION Continental and local declines in numerous bird populations have lead to concern for the future of migratory and resident bird species. The reasons for declines are complex. Habitat loss, modification, and fragmentation, loss of wintering and migratory habitat, and brood parasitism have been implicated. Scientists and the concerned public agreed that they needed a coordinated, cooperative conservation initiative focusing on birds. Partners in Flight was conceived as a voluntary, international coalition of government agencies, conservation groups, academic institutions, private businesses, and everyday citizens dedicated to “keeping common birds common.” PURPOSE AND SCOPE Idaho Partners in Flight focused this plan on restoring healthy ecosystems that will maintain productive and complete bird communities. The plan identifies priority bird species, then uses those species and other information on habitat trends to focus on the highest priority habitats. Thus, this plan takes a habitat-based approach, rather than a species-based approach, to conserving bird populations. This version of the Idaho Bird Conservation Plan covers in detail four habitats that we consider the highest priority habitats for birds in Idaho: Riparian; Non-riverine Wetlands; Sagebrush Shrublands; and Dry Ponderosa Pine/Douglas-fir/Grand Fir Forests. For each of these habitats, we describe their importance to birds, give habitat descriptions, state objectives and issues, and give strategies and tasks for meeting those objectives. In future versions of the Idaho Bird Conservation Plan, we will address other habitats important to birds.