Bison Conservation in the Northern Great Plains (2008)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

AMERICAN PRAIRIE RESERVE SEES RECORD VISITATION for 2020 Lodging Reservation System Now Open for 2021
FOR IMMEDIATE RELEASE FEBRUARY 1, 2021 Contact: Beth Saboe Senior Media and Government Relations Manager Phone: 406-600-4906 Email: [email protected] AMERICAN PRAIRIE RESERVE SEES RECORD VISITATION FOR 2020 Lodging reservation system now open for 2021 American Prairie Reserve experienced a surge of visitation in 2020, recording more people staying overnight at its facilities than any previous year. Overnight reservations last year were up nearly 200 percent compared to 2019. The uptick in visitation was driven in part by people seeking respite outdoors from the ongoing coronavirus pandemic, and came despite having only 40 percent of the Reserve’s camping and lodging inventory available due to COVID-19 precautions. According to American Prairie’s Director of Recreation and Public Access Mike Kautz, 665 reservations were recorded in 2020 from the Reserve’s three huts, and two campgrounds. In comparison, there were 232 reservations recorded in 2019. A reservation is defined as a group reserving one unit of lodging (campsite, hut or cabin), and not a count of individual visitors. Kautz says each reservation had an average group size of 4 people. In addition, the majority of people visiting the Reserve are Montanans. In 2020, more than 88% of the reservations for American Prairie’s hut system and more than 50% of reservations for the campgrounds came from state residents. Kautz says this growing interest suggests visitors should consider making reservations earlier this year. “Word of this pretty unique prairie experience we offer is quickly spreading,” said Kautz. “We fully expect our reservation inventory to fill up quickly this year so anyone who is curious about the lodging options shouldn’t wait too long.” Reservations for American Prairie’s 2021 visitation season are now being accepted, as of February 1. -
National Register of Historic Places Faster Registration Form
/ & NFS Form 10-900 OMB No. 1024-0018 (Rev. 8-86) United States Department of the Interior - - National Park Service - •- -' f\VM;OMAl NATIONAL REGISTER OF HISTORIC PLACES FASTER REGISTRATION FORM 1. Name of Property historic name: Hornaday Camp other name/site number: 246F362 2. Location street & number: Montana Highway 200 not for publication: n/a vicinity: n/a city/town: Sand Springs state: Montana code: MT county: Gar field code: 033 zip code: 59077 3. Classification Ownership of Property: private Category of Property: site Number of Resources within Property: Contributing Noncontributing ____ ____ building(s) 1 ____ sites ____ ____ structures ____ ____ objects Total Number of contributing resources previously listed in the National Register: 0 Name of related multiple property listing: n/a 4. Certification As the designated authority under the National Historic Preservation Act of 1986, as amended, I hereby certify that this X nomination ___request for determination of eligibility meets the documentation standards for registering properties in the National Register of Historic Places and meets the procedural and professional requirements set forth in 36 CFR Part 60. In my opinion, the property X meets ___ does not meet the National Register Criteria. ____ See continuation sheet. Signature of certifying official U 0 Date 0 State or Federal agency and bureau In my opinion, the property ___ meets ___ does not meet the National Register criteria. __ See continuation sheet. Signature of commenting or other official Date State or Federal agency and bureau 5. National Park Service Certification I, hereby certify that this property is: v/ entered in the National Register (LuJfflWfftyjL,(I ' MU.hD __ See continuation sheet. -

Bison Literature Review Biology
Bison Literature Review Ben Baldwin and Kody Menghini The purpose of this document is to compare the biology, ecology and basic behavior of cattle and bison for a management context. The literature related to bison is extensive and broad in scope covering the full continuum of domestication. The information incorporated in this review is focused on bison in more or less “wild” or free-ranging situations i.e.., not bison in close confinement or commercial production. While the scientific literature provides a solid basis for much of the basic biology and ecology, there is a wealth of information related to management implications and guidelines that is not captured. Much of the current information related to bison management, behavior (especially social organization) and practical knowledge is available through local experts, current research that has yet to be published, or popular literature. These sources, while harder to find and usually more localized in scope, provide crucial information pertaining to bison management. Biology Diet Composition Bison evolutional history provides the basis for many of the differences between bison and cattle. Bison due to their evolution in North America ecosystems are better adapted than introduced cattle, especially in grass dominated systems such as prairies. Many of these areas historically had relatively low quality forage. Bison are capable of more efficient digestion of low-quality forage then cattle (Peden et al. 1973; Plumb and Dodd 1993). Peden et al. (1973) also found that bison could consume greater quantities of low protein and poor quality forage then cattle. Bison and cattle have significant dietary overlap, but there are slight differences as well. -

5Th American Bison Society Meeting and Workshop
5th American Bison Society Meeting and Workshop Banff, Alberta September 26-29, 2016 Cover photos: Bison photos: © Kent Redford; Treaty signing: © Stephen Legault h Message from the Governor General I am delighted to extend my warmest regards to all those gathered for the 2016 American Bison Society Meeting. Over a century ago, long before the advent of ‘green’ living, a passionate group of individuals banded together to revitalize the dwindling North American bison population. These magnificent animals had been ravaged by human greed, almost to the point of extinction. Yet, through the efforts of the American Bison Society, the bison have returned to the wild in great numbers. The Society continues to play an important role in ensuring the survival of the bison. Its members have adopted the values of preservation and conservation, and are sharing this knowledge with the next generation, so that they may carry on this essential task. I commend the Society on its achievements and I wish everyone a most enjoyable celebration. David Johnston September 2016 5th American Bison Society Meeting and Workshop H American Bison Society h Welcome from WCS Dear colleagues, Welcome to the fifth bi-annual American Bison Society meeting and workshop. For the first time, we are hosting this meeting of bison enthusiasts, managers, producers, advocates, philanthropists, and artists in Canada. Not only is Banff a beautiful setting, it also plays a crucial role in the history of bison in North America. Over the next three days we will come together to share stories, learn about our work across North America and, finally, celebrate the second anniversary of the Buffalo Treaty. -

Central Mixedgrass Prairie Ecological System (Central Shortgrass Prairie Ecoregion Version)
CENTRAL MIXEDGRASS PRAIRIE ECOLOGICAL SYSTEM (CENTRAL SHORTGRASS PRAIRIE ECOREGION VERSION) ECOLOGICAL INTEGRITY ASSESSMENT Draft of June 29, 2007 Prepared by: Karin Decker Colorado Natural Heritage Program Colorado State University 254 General Services Building Fort Collins, CO 80523 Table of Contents A. INTRODUCTION ........................................................................................................ 3 A.1 Classification Summary ........................................................................................... 3 A.2 Ecological System Description ................................................................................ 5 A.2.1 Environment....................................................................................................... 5 A.2.2 Vegetation & Ecosystem.................................................................................... 6 A.2.3 Dynamics ........................................................................................................... 8 A.2.4 Landscape......................................................................................................... 10 A.2.5 Size................................................................................................................... 11 A.3 Ecological Integrity................................................................................................ 12 A.3.1 Threats.............................................................................................................. 12 A.3.2 Justification of Metrics.................................................................................... -

Buffalo Hunt: International Trade and the Virtual Extinction of the North American Bison
NBER WORKING PAPER SERIES BUFFALO HUNT: INTERNATIONAL TRADE AND THE VIRTUAL EXTINCTION OF THE NORTH AMERICAN BISON M. Scott Taylor Working Paper 12969 http://www.nber.org/papers/w12969 NATIONAL BUREAU OF ECONOMIC RESEARCH 1050 Massachusetts Avenue Cambridge, MA 02138 March 2007 I am grateful to seminar participants at the University of British Columbia, the University of Calgary, the Environmental Economics workshop at the NBER Summer Institute 2006, the fall 2006 meetings of the NBER ITI group, and participants at the SURED II conference in Ascona Switzerland. Thanks also to Chris Auld, Ed Barbier, John Boyce, Ann Carlos, Charlie Kolstad, Herb Emery, Mukesh Eswaran, Francisco Gonzalez, Keith Head, Frank Lewis, Mike McKee, and Sjak Smulders for comments; to Michael Ferrantino for access to the International Trade Commission's library; and to Margarita Gres, Amanda McKee, Jeffrey Swartz, Judy Hasse of Buffalo Horn Ranch and Andy Strangeman of Investra Ltd. for research assistance. Funding for this research was provided by the SSHRC. The views expressed herein are those of the author(s) and do not necessarily reflect the views of the National Bureau of Economic Research. © 2007 by M. Scott Taylor. All rights reserved. Short sections of text, not to exceed two paragraphs, may be quoted without explicit permission provided that full credit, including © notice, is given to the source. Buffalo Hunt: International Trade and the Virtual Extinction of the North American Bison M. Scott Taylor NBER Working Paper No. 12969 March 2007 JEL No. F1,Q2,Q5,Q56 ABSTRACT In the 16th century, North America contained 25-30 million buffalo; by the late 19th century less than 100 remained. -

Northern Mixed-Grass Prairie Is Strongly Influenced by Wet-Dry Cycles
Rapid Assessment Reference Condition Model The Rapid Assessment is a component of the LANDFIRE project. Reference condition models for the Rapid Assessment were created through a series of expert workshops and a peer-review process in 2004 and 2005. For more information, please visit www.landfire.gov. Please direct questions to [email protected]. Potential Natural Vegetation Group (PNVG) R4PRMGn Northern Mixed Grass Prairie General Information Contributors (additional contributors may be listed under "Model Evolution and Comments") Modelers Reviewers Cody Wienk [email protected] David Engle [email protected] Lakhdar Benkobi [email protected] John Ortmann [email protected] Vegetation Type General Model Sources Rapid AssessmentModel Zones Grassland Literature California Pacific Northwest Local Data Great Basin South Central Dominant Species* Expert Estimate Great Lakes Southeast Northeast S. Appalachians AGSM KOMA LANDFIRE Mapping Zones STIPA BUDA Northern Plains Southwest 30 39 N-Cent.Rockies BOUT 31 40 CAFI 33 Geographic Range Northeastern Montana, western North and South Dakota, northeastern Wyoming, western Nebraska. Biophysical Site Description Elevations range from 1,300 to 4,000 feet. Temperatures range between extremes of hot summers and cold winters that are typical of a continental climate. Precipitation increases from west (12 in.) to east (24 in.). Two-thirds of the precipitation occurs during the growing season. Soils vary, but are generally aridicols in the west and mollisols in the east. Soils in the northern Great Plains, west of the Missouri River in the Dakotas, northwestern Nebraska, northeastern Wyoming and Montana are formed from sandstone and shales. These soils range from clayey, fine-loamy, to fine silty soils of mixed origin in level and hilly- undulating lands with major contributions from loess, eolian sand, alluvium, and mountain outwash. -

Last Interglacial (MIS 5) Ungulate Assemblage from the Central Iberian Peninsula: the Camino Cave (Pinilla Del Valle, Madrid, Spain)
Palaeogeography, Palaeoclimatology, Palaeoecology 374 (2013) 327–337 Contents lists available at SciVerse ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Last Interglacial (MIS 5) ungulate assemblage from the Central Iberian Peninsula: The Camino Cave (Pinilla del Valle, Madrid, Spain) Diego J. Álvarez-Lao a,⁎, Juan L. Arsuaga b,c, Enrique Baquedano d, Alfredo Pérez-González e a Área de Paleontología, Departamento de Geología, Universidad de Oviedo, C/Jesús Arias de Velasco, s/n, 33005 Oviedo, Spain b Centro Mixto UCM-ISCIII de Evolución y Comportamiento Humanos, C/Sinesio Delgado, 4, 28029 Madrid, Spain c Departamento de Paleontología, Facultad de Ciencias Geológicas, Universidad Complutense de Madrid, Ciudad Universitaria, 28040 Madrid, Spain d Museo Arqueológico Regional de la Comunidad de Madrid, Plaza de las Bernardas, s/n, 28801-Alcalá de Henares, Madrid, Spain e Centro Nacional de Investigación sobre la Evolución Humana (CENIEH), Paseo Sierra de Atapuerca, s/n, 09002 Burgos, Spain article info abstract Article history: The fossil assemblage from the Camino Cave, corresponding to the late MIS 5, constitutes a key record to un- Received 2 November 2012 derstand the faunal composition of Central Iberia during the last Interglacial. Moreover, the largest Iberian Received in revised form 21 January 2013 fallow deer fossil population was recovered here. Other ungulate species present at this assemblage include Accepted 31 January 2013 red deer, roe deer, aurochs, chamois, wild boar, horse and steppe rhinoceros; carnivores and Neanderthals Available online 13 February 2013 are also present. The origin of the accumulation has been interpreted as a hyena den. Abundant fallow deer skeletal elements allowed to statistically compare the Camino Cave fossils with other Keywords: Early Late Pleistocene Pleistocene and Holocene European populations. -

COMMUNITY CONSERVATION PLAN Southwestern Manitoba Mixed
Southwestern Manitoba Mixed-grass Prairie IBA Page 1 of 1 COMMUNITY CONSERVATION PLAN for the Southwestern Manitoba Mixed-grass Prairie IMPORTANT BIRD AREA A Grassland Bird Initiative for Southwestern Manitoba's - • Poverty Plains • Lyleton-Pierson Prairies • Souris River Lowlands Prepared by: Cory Lindgren, Ken De Smet Manitoba IBA Program Species At Risk Biologist Oak Hammock Marsh Wildlife Branch, Manitoba Conservation Box 1160, Stonewall, Manitoba R0E 2Z0 200 Saulteaux Crescent, Winnipeg R3J 3W3 Manitoba IBA Program 10/03/01 _____________________________________________________________________________________ Southwestern Manitoba Mixed-grass Prairie IBA Page 2 of 2 Table of Contents 1.0 INTRODUCTION .................................................................................................................................. 8 1.1 The Poverty Plains.......................................................................................................................... 8 1.2 Souris River Lowlands ................................................................................................................... 8 1.3 Lyleton-Pierson Prairies ................................................................................................................ 9 2.0 THE IBA PROGRAM........................................................................................................................... 9 2.1 IBA Manitoba ........................................................................................................................... -

A Middle Holocene Steppe Bison and Paleoenvironments from the Versleuce Meadows, Whitehorse, Yukon, Canada Grant D
1138 ARTICLE A middle Holocene steppe bison and paleoenvironments from the Versleuce Meadows, Whitehorse, Yukon, Canada Grant D. Zazula, Elizabeth Hall, P. Gregory Hare, Christian Thomas, Rolf Mathewes, Catherine La Farge, André L. Martel, Peter D. Heintzman, and Beth Shapiro Abstract: A partial skeleton of a bison was recovered during residential house construction in Whitehorse, Yukon, Canada. The specimen represents a young (estimated 6 year old) bison individual that died, was partially scavenged by carnivores, and subsequently buried by calcareous silt sediment in a pond or small lake during the middle Holocene, ϳ5400 years ago. Palaeoenvironmental data, including molluscs, pollen, vascular plant, and bryophyte macrofossils demonstrate that the small waterbody was surrounded by white spruce dominated boreal forest. Morphometric analysis of the skeleton reveals that its taxonomic affinity is ambiguous, likely owing to it representing an ontogenetically young individual, though it does share some cranial and horn core characteristics of named species such as Bison occidentalis or Bison priscus. Mitochondrial genomic data confirm that this bison belongs to Clade 2A (northern clade), which represents Pleistocene steppe bison (B. cf. priscus) in Beringia through the Holocene and is not represented in living bison species. These data further demonstrate that northern steppe bison population survived the late Pleistocene extinction event, persisted locally in southern Yukon into the Holocene, and are best characterized as a species with a high degree of morphological variability and ecological flexibility. Résumé : Le squelette partiel d’un bison a été récupéré durant la construction d’immeubles résidentiels a` Whitehorse (Yukon, Canada). Le spécimen représente un unique jeune bison (d’âge estimé a` 6 ans) qui est mort, a été partiellement dévoré par des carnivores pour ensuite être enseveli sous du silt calcareux dans un étang ou un petit lac durant l’Holocène moyen, il y a ϳ5400 ans. -
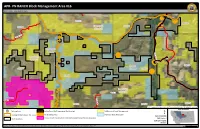
APR- PN RANCH Block Management Area #16 BMA Rules - See Reverse Page
APR- PN RANCH Block Management Area #16 BMA Rules - See Reverse Page Ch ip C re e k 23N15E 23N17E 23N16E !j 23N14E Deer & Elk HD # 690 Rd ille cev Gra k Flat Cree Missouri River k Chouteau e e r C County P g n o D B r i d g e Deer & Elk R d HD # 471 UV236 !j Fergus 22N17E 22N14E County 22N16E 22N15E A Deer & Elk r ro w The Peak C HD # 426 r e e k E v Judi e th r Riv s e ge Rd r Ran o n R d 21N17E 21N15E 21N16E U.S. Department of Agriculture Farm Services Agency Aerial Photography Field Office Area of Interest !j Parking Area Safety Zone (No Trespassing, No Hunting) US Bureau of Land Management º 1 6 Hunting Districts (Deer, Elk, Lion) No Shooting Area Montana State Trust Land 4 Date: 5/22/2020 2 Moline Ranch Conservation Easement (Seperate Permission Required) FWP Region 4 7 BMA Boundary 5 BLM 100K Map(s): 3 Winifred Possession of this map does not constitute legal access to private land enrolled in the BMA Program. This map may not depict current property ownership outside the BMA. It is every hunter's responsibility to know the 0 0.5 1 land ownership of the area he or she intends to hunt, the hunting regulations, and any land use restrictions that may apply. Check the FWP Hunt Planner for updates: http://fwp.mt.gov/gis/maps/huntPlanner/ Miles AMERICAN PRAIRIE RESERVE– PN RANCH BMA # 16 Deer/Elk Hunting District: 426/471 Antelope Hunting District: 480/471 Hunting Access Dates: September 1 - January 1 GENERAL INFORMATION HOW TO GET THERE BMA Type Acres County Ownership » North on US-191 15.0 mi 2 20,203 Fergus Private » North on Winifred Highway 24.0 mi » West on Main St. -

Riparian Restoration Summary
American Prairie Reserve & Prairie Stream Restoration Introduction to American Prairie Foundation American Prairie Foundation (APF) is spearheading a historic effort to reignite America’s passion for large-scale conservation by assembling the largest wildlife reserve of any kind in the lower 48 states. APF’s lands, called American Prairie Reserve (APR), are located on Montana’s Great Plains, just north of the Charles M. Russell National Wildlife Refuge and the Missouri River. Scientists have identified this place as an ideal location for the construction of a thriving prairie reserve capable of hosting an astonishing array of wildlife in an ecologically diverse environment. APF currently holds 123,346 acres of deeded and leased land. APF has three goals informing all activities on the Reserve: 1. Conservation: APF seeks to accumulate and manage enough private land that, when combined with existing public land, will create a fully functioning prairie-based wildlife reserve. When complete, the Reserve will consist of more than three million acres of private and public land (using the existing 1.1 million acre Charles M. Russell National Wildlife Refuge as the public land anchor). With less than 1% of the world’s grasslands protected, the pristine condition of the land and abundance of indigenous species in the Northern Great Plains make the American Prairie Reserve region an ideal location for a conservation project of this scale. APF currently manages a conservation bison herd of approximately 140 animals—the first conservation bison herd in this part of Montana in more than 120 years. Collaborative research projects are also underway to assess existing populations of endemic species including pronghorn, prairie dogs, swift fox, long- billed curlews, and cougars.