A Potential Target for the Treatment of Various Diseases
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

9-THC Disrupts Gamma (Γ)&Ndash;Band Neural
UCSF UC San Francisco Previously Published Works Title Δ9-THC Disrupts Gamma (γ)-Band Neural Oscillations in Humans Permalink https://escholarship.org/uc/item/6ms753gw Journal Neuropsychopharmacology, 40(9) ISSN 0893-133X Authors Cortes-Briones, J Skosnik, PD Mathalon, D et al. Publication Date 2015-08-16 DOI 10.1038/npp.2015.53 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Accepted Article Preview: Published ahead of advance online publication D9-THC Disrupts Gamma (c)–Band Neural Oscillations in Neuropsychopharmacology www.neuropsychopharmacology.org Humans Jose Cortes-Briones, Patrick D Skosnik, Daniel Mathalon, Methylphenidate modifies the motion of the circadian clock Lamotrigine in mood disorders and cocaine dependence John Cahill, Brian Pittman, Ashley Williams, R Andrew Cortical glutamate in postpartum depression Sewell, Mohini Ranganathan, Brian Roach, Judith Ford, Deepak Cyril D’Souza Cite this article as: Jose Cortes-Briones, Patrick D Skosnik, Daniel Mathalon, John Cahill, Brian Pittman, Ashley Williams, R Andrew Sewell, Mohini Ranganathan, Brian Roach, Judith Ford, Deepak Cyril D’Souza, D9-THC Disrupts Gamma (g)–Band Neural Oscillations in Humans, Neuropsychopharma- cology accepted article preview 24 February 2015; doi: 10.1038/npp.2015.53. This is a PDF file of an unedited peer-reviewed manuscript that has been accepted for publication. NPG are providing this early version of the manuscript as a service to our customers. The manuscript will undergo copyediting, typesetting and a proof review before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers apply. -

The Role of Organic Small Molecules in Pain Management
molecules Review The Role of Organic Small Molecules in Pain Management Sebastián A. Cuesta and Lorena Meneses * Laboratorio de Química Computacional, Facultad de Ciencias Exactas y Naturales, Escuela de Ciencias Químicas, Pontificia Universidad Católica del Ecuador, Av. 12 de Octubre 1076 Apartado, Quito 17-01-2184, Ecuador; [email protected] * Correspondence: [email protected]; Tel.: +593-2-2991700 (ext. 1854) Abstract: In this review, a timeline starting at the willow bark and ending in the latest discoveries of analgesic and anti-inflammatory drugs will be discussed. Furthermore, the chemical features of the different small organic molecules that have been used in pain management will be studied. Then, the mechanism of different types of pain will be assessed, including neuropathic pain, inflammatory pain, and the relationship found between oxidative stress and pain. This will include obtaining insights into the cyclooxygenase action mechanism of nonsteroidal anti-inflammatory drugs (NSAID) such as ibuprofen and etoricoxib and the structural difference between the two cyclooxygenase isoforms leading to a selective inhibition, the action mechanism of pregabalin and its use in chronic neuropathic pain, new theories and studies on the analgesic action mechanism of paracetamol and how changes in its structure can lead to better characteristics of this drug, and cannabinoid action mechanism in managing pain through a cannabinoid receptor mechanism. Finally, an overview of the different approaches science is taking to develop more efficient molecules for pain treatment will be presented. Keywords: anti-inflammatory drugs; QSAR; pain management; cyclooxygenase; multitarget drug; Citation: Cuesta, S.A.; Meneses, L. cannabinoid; neuropathic pain The Role of Organic Small Molecules in Pain Management. -

Edinburgh Research Explorer
Edinburgh Research Explorer International Union of Basic and Clinical Pharmacology. LXXXVIII. G protein-coupled receptor list Citation for published version: Davenport, AP, Alexander, SPH, Sharman, JL, Pawson, AJ, Benson, HE, Monaghan, AE, Liew, WC, Mpamhanga, CP, Bonner, TI, Neubig, RR, Pin, JP, Spedding, M & Harmar, AJ 2013, 'International Union of Basic and Clinical Pharmacology. LXXXVIII. G protein-coupled receptor list: recommendations for new pairings with cognate ligands', Pharmacological reviews, vol. 65, no. 3, pp. 967-86. https://doi.org/10.1124/pr.112.007179 Digital Object Identifier (DOI): 10.1124/pr.112.007179 Link: Link to publication record in Edinburgh Research Explorer Document Version: Publisher's PDF, also known as Version of record Published In: Pharmacological reviews Publisher Rights Statement: U.S. Government work not protected by U.S. copyright General rights Copyright for the publications made accessible via the Edinburgh Research Explorer is retained by the author(s) and / or other copyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associated with these rights. Take down policy The University of Edinburgh has made every reasonable effort to ensure that Edinburgh Research Explorer content complies with UK legislation. If you believe that the public display of this file breaches copyright please contact [email protected] providing details, and we will remove access to the work immediately and investigate your claim. Download date: 02. Oct. 2021 1521-0081/65/3/967–986$25.00 http://dx.doi.org/10.1124/pr.112.007179 PHARMACOLOGICAL REVIEWS Pharmacol Rev 65:967–986, July 2013 U.S. -

A Computational Approach for Defining a Signature of Β-Cell Golgi Stress in Diabetes Mellitus
Page 1 of 781 Diabetes A Computational Approach for Defining a Signature of β-Cell Golgi Stress in Diabetes Mellitus Robert N. Bone1,6,7, Olufunmilola Oyebamiji2, Sayali Talware2, Sharmila Selvaraj2, Preethi Krishnan3,6, Farooq Syed1,6,7, Huanmei Wu2, Carmella Evans-Molina 1,3,4,5,6,7,8* Departments of 1Pediatrics, 3Medicine, 4Anatomy, Cell Biology & Physiology, 5Biochemistry & Molecular Biology, the 6Center for Diabetes & Metabolic Diseases, and the 7Herman B. Wells Center for Pediatric Research, Indiana University School of Medicine, Indianapolis, IN 46202; 2Department of BioHealth Informatics, Indiana University-Purdue University Indianapolis, Indianapolis, IN, 46202; 8Roudebush VA Medical Center, Indianapolis, IN 46202. *Corresponding Author(s): Carmella Evans-Molina, MD, PhD ([email protected]) Indiana University School of Medicine, 635 Barnhill Drive, MS 2031A, Indianapolis, IN 46202, Telephone: (317) 274-4145, Fax (317) 274-4107 Running Title: Golgi Stress Response in Diabetes Word Count: 4358 Number of Figures: 6 Keywords: Golgi apparatus stress, Islets, β cell, Type 1 diabetes, Type 2 diabetes 1 Diabetes Publish Ahead of Print, published online August 20, 2020 Diabetes Page 2 of 781 ABSTRACT The Golgi apparatus (GA) is an important site of insulin processing and granule maturation, but whether GA organelle dysfunction and GA stress are present in the diabetic β-cell has not been tested. We utilized an informatics-based approach to develop a transcriptional signature of β-cell GA stress using existing RNA sequencing and microarray datasets generated using human islets from donors with diabetes and islets where type 1(T1D) and type 2 diabetes (T2D) had been modeled ex vivo. To narrow our results to GA-specific genes, we applied a filter set of 1,030 genes accepted as GA associated. -

Cannabinoid As Potential Aromatase Inhibitor Through Molecular Modeling and Screening for Anti-Cancer Activity
Cannabinoid as Potential Aromatase Inhibitor Through Molecular Modeling and Screening for Anti-Cancer Activity Sudipta Baroi1, Achintya Saha2, Ritesh Bachar3 and Sitesh C Bachar4 1Department of Pharmaceutical Technology, Faculty of Pharmacy, University of Dhaka Dhaka-1000, Bangladesh 2Department of Chemical Technology, Pharmaceutical & Fine Chemical Technology Division University of Calcutta, India 3Department of Pharmacy, School of Science and Engineering, University of Information Technology and Sciences (UITS), Dhaka-1212, Bangladesh 4Department of Pharmacy, Faculty of Pharmacy, University of Dhaka, Dhaka-1000, Bangladesh (Received: April 15, 2020; Accepted: June 2, 2020; Published (web): June 28, 2020) ABSTRACT: Inhibition of aromatase (CYTP450), a key enzyme in the estrogen biosynthesis, could result in regression of estrogen-dependent tumors and even prevent the promotion of breast cancer. The present research has been designed for searching a potent chemical moiety from natural sources to inhibit aromatase enzyme, the over- functionality of which causes the breast cancer. Cannabis sativa contains a very much promising group of cannabinoids with more than 66 compounds with reported anticancer property and for the search of a target specific potent aromatase inhibitor, 61 cannabinoids from C. sativa were selected. The Structures Data File (SDF) of these ligand molecules were subjected to docking studies at the binding site of aromatase X-ray crystallographic structure based on lower resolution of the protein crystal structure and higher docking accuracy, predicted by calculating the correlation between experimental activities and Glide dock scores and compared with the standard aromatase ligand androstenedione and aromatase inhibitor fadrozole with existing drug for breast cancer treatment. The best docked pose of each ligand was selected on the basis of the highest dock score related to the binding free energies of the internal dataset compounds as compared to their observed activities. -

Compendium of Drug Abuse Jargon
Compendium of Drug Abuse Jargon Ronald P. Evens, PharmD William Clementi, BS (Pharm) San Antonio and Austin, Texas Drug abuse slang is an originative and a protean language, which poses the problem of identification and definition of terms. This compilation of nomenclature provides an extensive list to serve as a resource for the busy family physician, who frequently deals with drug abuse problems. Despite efforts by the government encompassing the following abuse The list on the following pages is and in the private sector to identify agents: alcohol, glue, hallucinogens, organized in alphabetical sequence drug abuse problems and collate the narcotics, sedative-hypnotics, and according to the drug abuse term. terminology of the “drug culture,” the stimulants; and the street slang em resultant lists of nomenclature usually ployed in association with their distri References describe only the “top 100 or 50” bution and administration, and 1. Konner DD, Rubin I: The pharma terms. This severely limits their useful reactions to them. cist's drug abuse manual. Pharmacy Times 39:35-37, 1973 ness as a resource tool because a The difficulty in comprehension of 2. Narcotics Identification Guide. significant proportion of the terms are this specialized lingo is primarily due Richmond, Va, Jump Street, The Drug Treatment Program of the Medical College already familiar to practitioners while to its protean nature and the continual of Virginia, 1970 generation of new terms. Different 3. Some Substances Used for Non- a vastly greater number of terms in use prescribed Drugging Effects. Washington, do not appear on the lists at all. subpopulations in the drug abuse cul DC, Pharmaceutical Manufacturers Associa tion, January, 1973 Therefore, we have accumulated and ture use dissimilar, changeable subsets 4. -

Identification of Biomarkers for Sarcoidosis and Tuberculosis of The
DATABASE ANALYSIS e-ISSN 1643-3750 © Med Sci Monit, 2020; 26: e925438 DOI: 10.12659/MSM.925438 Received: 2020.04.26 Accepted: 2020.05.11 Identification of Biomarkers for Sarcoidosis and Available online: 2020.05.28 Published: 2020.07.23 Tuberculosis of the Lung Using Systematic and Integrated Analysis Authors’ Contribution: ABCEF 1 Min Zhao 1 Department of Respiratory and Critical Care Medicine, The Second Hospital of Study Design A BCE 1 Xin Di Jilin University, Changchun, Jilin, P.R. China Data Collection B 2 Department of Oncology and Hematology, The Second Hospital of Jilin University, Statistical Analysis C CDE 2 Xin Jin Changchun, Jilin, P.R. China Data Interpretation D BF 1 Chang Tian Manuscript Preparation E CF 1 Shan Cong Literature Search F Funds Collection G BF 1 Jiangyin Liu ABDEG 1 Ke Wang Corresponding Author: Ke Wang, e-mail: [email protected] Source of support: This study was supported by the Technology Research Funds of Jilin Province (20190303162SF), the Natural Science Foundation of Jilin Province (20180101103JC), and the Medical and Health Project Funds of Jilin Province (20191102012YY) Background: Sarcoidosis (SARC) is a multisystem inflammatory disease of unknown etiology and pulmonary tuberculosis (PTB) is caused by Mycobacterium tuberculosis. Both of these diseases affect lungs and lymph nodes and share similar clinical manifestations. However, the underlying mechanisms for the similarities and differences in ge- netic characteristics of SARC and PTB remain unclear. Material/Methods: Three datasets (GSE16538, GSE20050, and GSE19314) were retrieved from the Gene Expression Omnibus (GEO) database. Differentially expressed genes (DEGs) in SARC and PTB were identified using GEO2R online analyz- er and Venn diagram software. -

NIH Public Access Author Manuscript Drug Alcohol Depend
NIH Public Access Author Manuscript Drug Alcohol Depend. Author manuscript; available in PMC 2015 October 01. NIH-PA Author ManuscriptPublished NIH-PA Author Manuscript in final edited NIH-PA Author Manuscript form as: Drug Alcohol Depend. 2014 October 1; 143: 251–256. doi:10.1016/j.drugalcdep.2014.08.004. Effects of the cannabinoid CB1 receptor allosteric modulator ORG 27569 on reinstatement of cocaine- and methamphetamine- seeking behavior in rats* Li Jinga,b, Yanyan Qiua, Yanan Zhangc, and Jun-Xu Lia aDepartment of Pharmacology and Toxicology, University at Buffalo, Buffalo, New York, USA bDepartment of Physiology and Pathophysiology, Tianjin Medical University, Tianjin, China cResearch Triangle Institute, Research Triangle Park, North Carolina, USA Abstract Background—Cannabinoid CB1 receptors play an essential role in drug addiction. Given the side effect profiles of orthosteric CB1 antagonism, new strategies have been attempted to modulate this target, such as CB1 receptor allosteric modulation. However, the effect of CB1 allosteric modulation in drug addiction is unknown. The present study examined the effects of the CB1 receptor allosteric modulator ORG27569 on the reinstatement of cocaine- and methamphetamine- seeking behavior in rats. Methods—Rats were trained to self-administer 0.75 mg/kg cocaine or 0.05 mg/kg methamphetamine in 2-hr daily sessions for 14 days which was followed by 7 days of extinction sessions in which rats responded on the levers with no programmed consequences. On reinstatement test sessions, rats were administered ORG27569 (1.0, 3.2, 5.6 mg/kg, i.p.) or SR141716A (3.2 mg/kg, i.p.) 10 min prior to re-exposure to cocaine- or methamphetamine-paired cues or a priming injection of cocaine (10 mg/kg, i.p.) or methamphetamine (1 mg/kg, i.p.). -

Pharmacy and Poisons (Third and Fourth Schedule Amendment) Order 2017
Q UO N T FA R U T A F E BERMUDA PHARMACY AND POISONS (THIRD AND FOURTH SCHEDULE AMENDMENT) ORDER 2017 BR 111 / 2017 The Minister responsible for health, in exercise of the power conferred by section 48A(1) of the Pharmacy and Poisons Act 1979, makes the following Order: Citation 1 This Order may be cited as the Pharmacy and Poisons (Third and Fourth Schedule Amendment) Order 2017. Repeals and replaces the Third and Fourth Schedule of the Pharmacy and Poisons Act 1979 2 The Third and Fourth Schedules to the Pharmacy and Poisons Act 1979 are repealed and replaced with— “THIRD SCHEDULE (Sections 25(6); 27(1))) DRUGS OBTAINABLE ONLY ON PRESCRIPTION EXCEPT WHERE SPECIFIED IN THE FOURTH SCHEDULE (PART I AND PART II) Note: The following annotations used in this Schedule have the following meanings: md (maximum dose) i.e. the maximum quantity of the substance contained in the amount of a medicinal product which is recommended to be taken or administered at any one time. 1 PHARMACY AND POISONS (THIRD AND FOURTH SCHEDULE AMENDMENT) ORDER 2017 mdd (maximum daily dose) i.e. the maximum quantity of the substance that is contained in the amount of a medicinal product which is recommended to be taken or administered in any period of 24 hours. mg milligram ms (maximum strength) i.e. either or, if so specified, both of the following: (a) the maximum quantity of the substance by weight or volume that is contained in the dosage unit of a medicinal product; or (b) the maximum percentage of the substance contained in a medicinal product calculated in terms of w/w, w/v, v/w, or v/v, as appropriate. -
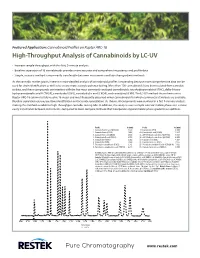
High-Throughput Analysis of Cannabinoids by LC-UV
Featured Application: Cannabinoid Profiles on Raptor ARC-18 High-Throughput Analysis of Cannabinoids by LC-UV • Increase sample throughput with this fast, 9-minute analysis. • Baseline separation of 16 cannabinoids provides more accurate and comprehensive potency and profile data. • Simple, isocratic method is more easily transferable between instruments and labs than gradient methods. As the cannabis market grows, interest in more detailed analysis of cannabinoid profiles is expanding because more comprehensive data can be used for strain identification as well as to ensure more accurate potency testing. More than 100 cannabinoids have been isolated from cannabis to date, and these compounds can interfere with the five most commonly analyzed cannabinoids: tetrahydrocannabinol (THC), delta-9-tetra- hydrocannabinolic acid A (THCA), cannabidiol (CBD), cannabidiolic acid (CBDA), and cannabinol (CBN). The LC-UV method shown here uses a Raptor ARC-18 column to fully resolve 16 major and most frequently observed minor cannabinoids for which commercial standards are available. Baseline separation ensures positive identification and accurate quantitation. As shown, all compounds were resolved in a fast 9-minute analysis, making this method suitable for high-throughput cannabis testing labs. In addition, this analysis uses a simple isocratic mobile phase so it is more easily transferable between instruments, compared to more complex methods that incorporate atypical mobile phase gradients or additives. Peaks tR (min) Peaks tR (min) 1. Cannabidivarinic acid (CBDVA) 1.877 9. Cannabinol (CBN) 4.609 2. Cannabidivarin (CBDV) 2.086 10. Cannabinolic acid (CBNA) 5.437 3. Cannabidiolic acid (CBDA) 2.592 11. Δ9-Tetrahydrocannabinol (Δ9-THC) 5.815 4. Cannabigerolic acid (CBGA) 2.750 12. -

CBD (Cannabidiol)
TRANSPORTATION RESEARCH BOARD Driving Toward the Truth - Dispelling the Myths About Cannabis Products February 10, 2021 @NASEMTRB #TRBwebinar PDH Certification The Transportation Research Board has met the standards and Information: requirements of the Registered Continuing Education Providers •1.5 Professional Development Program. Credit earned on completion Hour (PDH) – see follow-up of this program will be reported to email for instructions RCEP. A certificate of completion will •You must attend the entire be issued to participants that have registered and attended the entire webinar to be eligible to receive session. As such, it does not include PDH credits content that may be deemed or •Questions? Contact Reggie construed to be an approval or Gillum at [email protected] endorsement by RCEP. #TRBwebinar Learning Objectives 1. Identify impacts of the Farm Bill on use of THC and CBD products 2. Describe the toxicology of THC and CBD products 3. Discuss how THC and CBD products affect driving performance and crash risk #TRBwebinar TRB Standing Committee on Impairment in Transportation (ACS50) TRB Webinar: Driving Toward the Truth - Dispelling the Myths About Cannabis Products Dr. Barry K. Logan Executive Director, Center for Forensic Science Research and Education (CFSRE); Senior Vice President of Forensic Sciences, and Chief Scientist at NMS Labs Michelle Peace, Ph.D. Associate Professor and PI, Laboratory for Forensic Toxicology Research Department of Forensic Science, Virginia Commonwealth University Dr. Darrin Grondel Vice President, -

Potential Health Benefits of Cannabis Extracts: a Review Maria Rosana Ramirez
International Journal of Chemical and Biomedical Science 2016; 2(1): 1-8 Published online March 1, 2016 (http://www.aascit.org/journal/ijcbs) Potential Health Benefits of Cannabis Extracts: A Review Maria Rosana Ramirez CITER-The National Scientific and Technical Research Council (CONICET), Argentine Government, Agency, Argentina Email address [email protected] Citation Maria Rosana Ramirez. Potential Health Benefits of Cannabis Extracts: A Review. International Journal of Chemical and Biomedical Science . Vol. 2, No. 1, 2016, pp. 1-8. Keywords Abstract Non-cannabinoids Compounds, A central tenet underlying the use of plant preparations is that herbs contain many Cannabis Extracts, bioactive compounds. Cannabis contains tetrahydrocannabinols (THC) a primary Bioactivity metabolite with reported psychotropic effects. Therefore, the presence of THC makes controversial the use of Cannabis to treat diseases by which their uses and applications were limited. The question then is: is it possible to use the extracts from Cannabis to treat the diseases related with it use in folk medicine? More recently, the synergistic Received: January 24, 2016 contributions of bioactive constituents have been scientifically demonstrated. We Revised: February 10, 2016 reviewed the literature concerning medical cannabis and its secondary metabolites, Accepted: February 12, 2016 including fraction and total extracts. Scientific evidence shows that secondary metabolites in cannabis may enhance the positive effects of THC a primary metabolite. Other chemical components (cannabinoid and non-cannabinoid) in cannabis or its extracts may reduce THC-induced anxiety, cholinergic deficits, and immunosuppression; which could increase its therapeutic potential. Particular attention will be placed on non- cannabinoid compounds interactions that could produce synergy with respect to treatment of pain, inflammation, epilepsy, fungal and bacterial infections.