Sex and Race Differences in the Relative Lengths of Metacarpals and Metatarsals in Human Skeletons
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Metatarsalgia
just the symptoms. What can I expect from treatment? With a proper diagnosis, and a well-rounded treatment plan including orthotics, the prog- nosis is excellent. With Sole Supports™ foot or- thotics you can expect either a dramatic loss The Truth About . of pain within the first weeks of use or a more gradual reduction of symptoms, depending Metatarsalgia on how long the problem has existed, normal body weight or how well you follow other ther- For more information and a apeutic regimens prescribed by your provider. professional consultation regarding Did you know that, with Sole whether Sole Supports may be Supports, metatarsal pads are rarely helpful for you, please contact the needed? following certified Sole Supports practitioner: What is it? Metatarsalgia is a term used to describe a pain- ful foot condition in the area just before the Arch Flattened small toes (more commonly referred to as the ball of the foot). The condition is characterized by pain and inflammation on the sole in the region of the metatarsal heads, which are the ends of the long bones in your foot. The joint This handout provides a general overview on this capsule or tendons may also be inflamed. topic and may not apply to everyone. To find out if this handout applies to you and to get more infor- mation on this subject, consult with your certified Sole Supports practitioner. Arch Restored The pain is generally aggravated by putting ments but your doctor is likely to recommend pressure off the metatarsals should also be pressure (as in walking) through the ball of a conservative approach first including: followed. -

Wound Classification
Wound Classification Presented by Dr. Karen Zulkowski, D.N.S., RN Montana State University Welcome! Thank you for joining this webinar about how to assess and measure a wound. 2 A Little About Myself… • Associate professor at Montana State University • Executive editor of the Journal of the World Council of Enterstomal Therapists (JWCET) and WCET International Ostomy Guidelines (2014) • Editorial board member of Ostomy Wound Management and Advances in Skin and Wound Care • Legal consultant • Former NPUAP board member 3 Today We Will Talk About • How to assess a wound • How to measure a wound Please make a note of your questions. Your Quality Improvement (QI) Specialists will follow up with you after this webinar to address them. 4 Assessing and Measuring Wounds • You completed a skin assessment and found a wound. • Now you need to determine what type of wound you found. • If it is a pressure ulcer, you need to determine the stage. 5 Assessing and Measuring Wounds This is important because— • Each type of wound has a different etiology. • Treatment may be very different. However— • Not all wounds are clear cut. • The cause may be multifactoral. 6 Types of Wounds • Vascular (arterial, venous, and mixed) • Neuropathic (diabetic) • Moisture-associated dermatitis • Skin tear • Pressure ulcer 7 Mixed Etiologies Many wounds have mixed etiologies. • There may be both venous and arterial insufficiency. • There may be diabetes and pressure characteristics. 8 Moisture-Associated Skin Damage • Also called perineal dermatitis, diaper rash, incontinence-associated dermatitis (often confused with pressure ulcers) • An inflammation of the skin in the perineal area, on and between the buttocks, into the skin folds, and down the inner thighs • Scaling of the skin with papule and vesicle formation: – These may open, with “weeping” of the skin, which exacerbates skin damage. -

Signs to Help the Deaf Included in This Packet
Signs to help the Deaf Included in this packet: Medical Signs Color Signs People Signs This is made by: Deanna Zander, I am a parent of a deaf son. Here is my email address: [email protected], if you have any questions, please email your question, Please put in the Subject box- RE: Medical Signs. For more information, or to obtain a hospital kit for Deaf or Hard of Hearing, please contact Pam Smith, Adult Outreach Coordinator @ 701-665-4401 Medical Signs (Medicine) Medical Medicine- Tip of bent middle finger rubs circle on left palm Sign- Palm-out indexes circle Signs alternately Made By: Deanna Zander Hi, Hello, Howdy Good-Bye, Yes, Yep Bye Right “S” hand & head nods (both head & hand nod) No, Nope Flat hand, Touch forehead, Just wave First two fingers close onto move forward slightly right thumb, & shake your head Appointment Schedule Fingertips of the right palm- out “5” draws down left palm; then turn palm-in & draws across palm The right “S” hand palm down, is postitioned above the left “S” hand, also palm- down. The right hand circels above the left in a clockwise manner & is brought on the back of left hand. Your Name, My Name Birthday My, Mine- Your- The right middle finger touches the chin, Palm of flat Vertical flat palm moves then moves down to touch the chest hand on chest toward person Fingerspell- The right hand, palm- out, is move left to right, fingers wiggling up & Name- Right “H” touches left “H” at right angles MM/DD/YYYY down Call, phone Left “Y” hand, thumb near ear, little finger near mouth The upturned thumbs -

PE2260 Five-Finger Exercise
The Five-Finger Exercise The 5-finger exercise is helpful for relaxation and calming your system. It does not take long, but can help you feel much more peaceful and relaxed and help you feel better about yourself. Try it any time you feel tension. What are the steps to the 5-finger exercise? On one hand, touch your thumb to your index finger. Think back to a time you felt tired after exercise or some other fun physical activity. Touch your thumb to your middle finger. Go back to a time when you had a loving experience. You might recall a loving day with your family or a good friend, a warm hug from a parent or a time you had a really good conversation with someone. Touch your thumb to your ring finger. Remember the nicest compliment anyone ever gave you. Try to accept it now fully. When you do this, you are showing respect for the person who said it. You are really paying them a compliment in return. Touch your thumb to your little finger. Go back in your mind to the most beautiful and relaxing place you have ever been. Spend some time thinking of being there. To Learn More Free Interpreter Services • Adolescent Medicine • In the hospital, ask your nurse. 206-987-2028 • From outside the hospital, call the toll-free Family Interpreting Line, • Ask your healthcare provider 1-866-583-1527. Tell the interpreter • seattlechildrens.org the name or extension you need. Seattle Children’s offers interpreter services for Deaf, hard of hearing or non-English speaking patients, family members and legal representatives free of charge. -

Birth Cont R Ol Fact Sheet
VAGINAL RING FACT SHEET What is the Vaginal Ring (Nuvaring®)? The Vaginal Ring is a clear, flexible, thin, plastic ring that you place in the vagina where it stays for one cycle providing a continuous low dose of 2 hormones (estrogen and progestin). It prevents pregnancy by stopping the release of an egg (ovulation), thickening the cervical fluid, and changing the lining of the uterus. How effective is the Vaginal Ring? The ring is a very effective method of birth control. The ring is about 93% effective at preventing pregnancy in typical use, which means that around 7 out of 100 people who use it as their only form of birth control will get pregnant in one year. With consistent and correct use as described in this fact sheet, it can be over 99% effective. How can I get the Vaginal Ring? You can visit a clinic to get the ring or a prescription for it and talk with a healthcare provider about whether the ring is right for you. Advantages of the Vaginal Ring Disadvantages of the Vaginal Ring Periods may be more predictable/regular and lighter Must remember to remove and replace the ring once a Less period cramping month Decreased symptoms of Premenstrual Syndrome Some users may experience mild side effects such as: (PMS) and perimenopause spotting, nausea, breast tenderness, headaches, or Can be used to skip or shorten your periods dizziness (usually these improve in the first few months Less anemia/iron deficiency caused by heavy periods of use) Does not affect your ability to get pregnant in the Possibility of high blood pressure -

Hair-Thread Tourniquet Syndrome in an Infant with Bony Erosion a Case Report, Literature Review, and Meta-Analysis
REVIEW ARTICLE Hair-Thread Tourniquet Syndrome in an Infant With Bony Erosion A Case Report, Literature Review, and Meta-analysis Arman Z. Mat Saad, MB, AFRCSI, Elizabeth M. Purcell, MB, and Jack J. McCann, FRCS(Plas) tissue to cause bony erosion of the underlying phalanx of a Abstract: Hair-thread tourniquet syndrome is a rare condition toe. where appendages are strangulated by an encircling strand of hair, a thread, or a fiber. The condition usually occurs in very young patients in the first few months of life. We present a unique case of CASE REPORT a 3-month-old baby girl with hair-thread tourniquet syndrome in A 3-month-old baby girl was referred to our unit from whom a hair cheese-wired through the skin and soft tissue of the toe the emergency department, with a history of irritability and a and caused bony erosion of the underlying phalanx. An extensive red swollen right middle (third) toe, which failed to resolve 3 literature review and meta-analysis of the topic are also presented. days after removal of a hair tourniquet (in the emergency department of the referring hospital). Key Words: hair, thread, toe, finger, penile, clitoris, tourniquet Careful examination in our own emergency department syndrome with loupe magnification showed intact skin on the toe and no (Ann Plast Surg 2006;57: 447–452) further evidence of a residual hair tourniquet. A course of antibiotics was prescribed for cellulitis, and improvement was noted on review in our outpatient department at 1 week. Six weeks later, the patient represented to the outpatient air-thread tourniquet syndrome is a rare condition that department, with recurrent swelling and redness of the toe. -

Hand Gestures
L2/16-308 More hand gestures To: UTC From: Peter Edberg, Emoji Subcommittee Date: 2016-10-31 Proposed characters Tier 1: Two often-requested signs (ILY, Shaka, ILY), and three to complete the finger-counting sets for 1-3 (North American and European system). None of these are known to have offensive connotations. HAND SIGN SHAKA ● Shaka sign ● ASL sign for letter ‘Y’ ● Can signify “Aloha spirit”, surfing, “hang loose” ● On Emojipedia top requests list, but requests have dropped off ● 90°-rotated version of CALL ME HAND, but EmojiXpress has received requests for SHAKA specifically, noting that CALL ME HAND does not fulfill need HAND SIGN ILY ● ASL sign for “I love you” (combines signs for I, L, Y), has moved into mainstream use ● On Emojipedia top requests list HAND WITH THUMB AND INDEX FINGER EXTENDED ● Finger-counting 2, European style ● ASL sign for letter ‘L’ ● Sign for “loser” ● In Montenegro, sign for the Liberal party ● In Philippines, sign used by supporters of Corazon Aquino ● See Wikipedia entry HAND WITH THUMB AND FIRST TWO FINGERS EXTENDED ● Finger-counting 3, European style ● UAE: Win, victory, love = work ethic, success, love of nation (see separate proposal L2/16-071, which is the source of the information below about this gesture, and also the source of the images at left) ● Representation for Ctrl-Alt-Del on Windows systems ● Serbian “три прста” (tri prsta), symbol of Serbian identity ● Germanic “Schwurhand”, sign for swearing an oath ● Indication in sports of successful 3-point shot (basketball), 3 successive goals (soccer), etc. HAND WITH FIRST THREE FINGERS EXTENDED ● Finger-counting 3, North American style ● ASL sign for letter ‘W’ ● Scout sign (Boy/Girl Scouts) is similar, has fingers together Tier 2: Complete the finger-counting sets for 4-5, plus some less-requested hand signs. -

Female Pelvic Relaxation
FEMALE PELVIC RELAXATION A Primer for Women with Pelvic Organ Prolapse Written by: ANDREW SIEGEL, M.D. An educational service provided by: BERGEN UROLOGICAL ASSOCIATES N.J. CENTER FOR PROSTATE CANCER & UROLOGY Andrew Siegel, M.D. • Martin Goldstein, M.D. Vincent Lanteri, M.D. • Michael Esposito, M.D. • Mutahar Ahmed, M.D. Gregory Lovallo, M.D. • Thomas Christiano, M.D. 255 Spring Valley Avenue Maywood, N.J. 07607 www.bergenurological.com www.roboticurology.com Table of Contents INTRODUCTION .................................................................1 WHY A UROLOGIST? ..........................................................2 PELVIC ANATOMY ..............................................................4 PROLAPSE URETHRA ....................................................................7 BLADDER .....................................................................7 RECTUM ......................................................................8 PERINEUM ..................................................................9 SMALL INTESTINE .....................................................9 VAGINAL VAULT .......................................................10 UTERUS .....................................................................11 EVALUATION OF PROLAPSE ............................................11 SURGICAL REPAIR OF PELVIC PROLAPSE .....................15 STRESS INCONTINENCE .........................................16 CYSTOCELE ..............................................................18 RECTOCELE/PERINEAL LAXITY .............................19 -
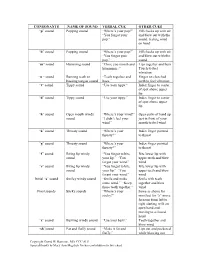
Consonants Name of Sound Verbal Cue Other Cues
CONSONANTS NAME OF SOUND VERBAL CUE OTHER CUES “p” sound Popping sound “Where’s your pop?” Fill cheeks up with air “You forgot your and blow out with the pop.” sound, feeling wind on hand “b” sound Popping sound “Where’s your pop?” Fill cheeks up with air “You forgot your and blow out with the pop.” sound “m” sound Humming sound “Close you mouth and Lips together and hum hummmm.:” Touch to feel vibration “n “ sound Buzzing teeth or “Teeth together and Finger on clenched buzzing tongue sound buzz.” teeth to feel vibration “t” sound Tippy sound “Use your tippy.” Index finger to center of spot above upper lip “d” sound Tippy sound “Use your tippy.” Index finger to center of spot above upper lip “h” sound Open mouth windy “Where’s your wind?” Open palm of hand up sound “I didn’t feel your just in front of your wind.” mouth to feel wind “k” sound Throaty sound “Where’s your Index finger pointed throaty?” to throat “g” sound Throaty sound “Where’s your Index finger pointed throaty?” to throat “f” sound Biting lip windy “You forgot to bite Bite lower lip with sound your lip.” “You upper teeth and blow forgot your wind.” wind “v” sound Biting lip windy “You forgot to bite Bite lower lip with sound your lip.” “You upper teeth and blow forgot your wind.” wind Initial “s” sound Smiley windy sound “Smile and make Smile with teeth some wind.” “Keep together and blow those teeth together.” wind Final sounds Sticky sounds “Where’s your Same as above for sticky?” most but for “s” move forearm form left to right starting with an open hand and moving to a closed hand “z” sound Buzzing windy sound “Use your buzz.” Teeth together and blow wind “sh”sound Fat and fluffy sound “Make it fat and Lips out and puckered fluffy.” while blowing out Copyright David W Hammer, MA CCC-SLP Special thanks to Mary Ann Migitsch for her contribution to this chart. -

Variations in the Finger Length of the Human Hand
Proceedings of the Iowa Academy of Science Volume 61 Annual Issue Article 63 1954 Variations in the Finger Length of the Human Hand Elizabeth Barnard Grinnell College G. Mendoza Grinnell College Let us know how access to this document benefits ouy Copyright ©1954 Iowa Academy of Science, Inc. Follow this and additional works at: https://scholarworks.uni.edu/pias Recommended Citation Barnard, Elizabeth and Mendoza, G. (1954) "Variations in the Finger Length of the Human Hand," Proceedings of the Iowa Academy of Science, 61(1), 458-462. Available at: https://scholarworks.uni.edu/pias/vol61/iss1/63 This Research is brought to you for free and open access by the Iowa Academy of Science at UNI ScholarWorks. It has been accepted for inclusion in Proceedings of the Iowa Academy of Science by an authorized editor of UNI ScholarWorks. For more information, please contact [email protected]. Barnard and Mendoza: Variations in the Finger Length of the Human Hand Variations in the Finger Length of the Human Hand By ELIZABETH BARNARD AND G. MENDOZA INTRODUCTION Although a great deal has been written concerning the occur rence of abnormalities of the hands and fingers, relatively few studies have been made to determine variations of the normal hand. The purpose of this study is to gather some valid statistics concerning the occurrence of variations in finger length within a segment of the general population. It is hoped that this study will serve as the beginning of a valid basis upon which a study of human inheritance can be built. Because the interindividual difference in the pattern of finger length consists in the relationship between the index and ring fingers, this varying relation has been most often reported in the literature. -

Cubital Tunnel Syndrome)
DISEASES & CONDITIONS Ulnar Nerve Entrapment at the Elbow (Cubital Tunnel Syndrome) Ulnar nerve entrapment occurs when the ulnar nerve in the arm becomes compressed or irritated. The ulnar nerve is one of the three main nerves in your arm. It travels from your neck down into your hand, and can be constricted in several places along the way, such as beneath the collarbone or at the wrist. The most common place for compression of the nerve is behind the inside part of the elbow. Ulnar nerve compression at the elbow is called "cubital tunnel syndrome." Numbness and tingling in the hand and fingers are common symptoms of cubital tunnel syndrome. In most cases, symptoms can be managed with conservative treatments like changes in activities and bracing. If conservative methods do not improve your symptoms, or if the nerve compression is causing muscle weakness or damage in your hand, your doctor may recommend surgery. This illustration of the bones in the shoulder, arm, and hand shows the path of the ulnar nerve. Reproduced from Mundanthanam GJ, Anderson RB, Day C: Ulnar nerve palsy. Orthopaedic Knowledge Online 2009. Accessed August 2011. Anatomy At the elbow, the ulnar nerve travels through a tunnel of tissue (the cubital tunnel) that runs under a bump of bone at the inside of your elbow. This bony bump is called the medial epicondyle. The spot where the nerve runs under the medial epicondyle is commonly referred to as the "funny bone." At the funny bone the nerve is close to your skin, and bumping it causes a shock-like feeling. -

Breathing Techniques for Kids (Toolkit)
BREATHING TECHNIQUES FOR KIDS exercises to center kids and help them focus CREATIVE TECHNIQUES SQUARE BREATHING On their desk or table, have kids trace a horizontal line with their fingers for a count of four as they breathe in (the top of the square). Then, trace downward to form the side of the square as they hold the breath for a count of four. Then they trace horizontally again to make the bottom of the square as they exhale. Finally, they trace upward to form the other side of the square as they hold their breath out for a count of four. Repeat. DRAW YOUR BREATH Give the children a marker and a sheet of paper. Have them place their marker on the paper. As they inhale and exhale, have them allow their markers to move up and down on the sheet. The end product is a scribble–an image of their breath! PHYSICAL TECHNIQUES TUMBLE DRYER Sitting in cross-legged position, point your index fingers towards each other and position them so your left finger is pointing to the right and your right finger is pointing to the left, overlapping a bit in front of your mouth. Inhale, then blow out as you spin your fingers round each other, making a long exhalation and a satisfying swishy sound. ALTERNATE NOSTRIL BREATHING For this breathing exercise, kids bring attention to their breath by holding one nostril closed as they breathe in and then holding the other nostril closed as they breathe out. SHOULDER ROLLS Sit comfortably. As you breathe in, roll your shoulders up and back.