Latitudinal Cline of Chronotype
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

406 | Tutto Arabi - Veterinary Biorhythm
406 | TUTTO ARABI - www.tuttoarabi.com VETERINARY BIORHYTHM by Domenico Bergero, Cynthia Préfontaine Rhythm! • infradian rhythms, which are long-term cycles, such as the To horses, as for men, the passing of time has an impact on annual migration or reproduction cycles found in certain many conscious or automatic activities. When it comes to animals or the human menstrual cycle. men, in particular athletes, we talk about “biorhythm”, i.e. the • ultradian rhythms, which are short cycles, such as the 90-minute effect that longer or shorter cycles can have on performance. REM cycle, the 4-hour nasal cycle, or the 3-hour cycle of But what about our adventure buddy? growth hormone production. They have periods of less than 24 hours. Chronobiology • tidal rhythms, commonly observed in marine life, which follow Chronobiology, from the Greek “chronos” (time) and the (roughly) 12-hour transition from high to low tide “biologia” (study of life), is a field of science that examines and back. periodic (cyclic) phenomena in living organisms and their adaptation to solar and lunar related rhythms. Hormones and physical performance Chronobiological studies include but are not limited to As everyone knows, physical performance is, among other comparative anatomy, physiology, genetics, molecular things, under the control of hormones that determine higher biology and behavior of organisms within biological rhythms or lesser fitness. These hormones can be produced continually mechanics. or, more often than not, can be mero present at certain The variations of the timing and duration of biological times (peak or acrophase) and then decrease. Let’s see which activity in living organisms occur for many essential hormones are involved and what are the best moments for biological processes. -

Latitudinal Cline of Chronotype
www.nature.com/scientificreports OPEN Latitudinal cline of chronotype Mario André Leocadio-Miguel 1, Fernando Mazzili Louzada2, Leandro Lourenção Duarte3, Roberta Peixoto Areas4, Marilene Alam5, Marcelo Ventura Freire4, John Fontenele-Araujo1, Luiz Menna-Barreto4 & Mario Pedrazzoli4 The rotation of the Earth around its own axis and around the sun determines the characteristics of Received: 15 February 2017 the light/dark cycle, the most stable and ancient 24 h temporal cue for all organisms. Due to the tilt in the earth’s axis in relation to the plane of the earth’s orbit around the sun, sunlight reaches the Earth Accepted: 2 June 2017 differentially depending on the latitude. The timing of circadian rhythms varies among individuals of Published: xx xx xxxx a given population and biological and environmental factors underlie this variability. In the present study, we tested the hypothesis that latitude is associated to the regulation of circadian rhythm in humans. We have studied chronotype profiles across latitudinal cline from around 0° to 32° South in Brazil in a sample of 12,884 volunteers living in the same time zone. The analysis of the results revealed that humans are sensitive to the different sunlight signals tied to differences in latitude, resulting in a morning to evening latitudinal cline of chronotypes towards higher latitudes. The concept of chronotype, which is the expression of diurnal preferences or circadian phenotype, including rise and bedtime preferences, has received a strong consideration from studies in human chronobiology1. It is generally accepted that the chronotype distribution in populations is the same, no matter where the population is geographically localized. -
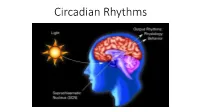
Circadian Rhythms Controlling the Timing of Behaviour by Anticipating the Environment
Circadian Rhythms Controlling the timing of behaviour by anticipating the environment • Circadian = circa + dium • Exists in most if not all unicellular and multicellular organisms The Circadian Circuit Environmental Output Inputs Rhythms Light Hormonal Cycles Temperature Rest/Wake Social Activity Feeding Central Pacemaker Health consequences of circadian misalignment Social Jet Lag Shift Work Increased risk of: - Obesity - Diabetes - Cancer - Mental Illness Light at Night Roenneberg et al. (2012) Current Biology Historical Perspective Jean-Jacques d’Ortous de Mairan (1678 – 1771) Hist de l’Acad Royal Sci (Paris), 1729 “…Il est seulement un peu moins marqué lorsqu’on la tient toujours enfermée dans un lieu obscur…” “The sensitive plant hence perceives the sun without seeing it” Rhythms in leaf-opening persist even in the absence of sunlight Historical Perspective Rat Comp Psychol Monographs, 1922 Nathaniel Kleitman (1895 – 1999) Figure 18.4 Sleep and Wakefulness, 1963 Historical Perspective ’Founders of Chronobiology’ 1960 • Conceptual framework of circadian rhythms • Long before any genes or neural circuits were identified Colin Pittendrigh Jürgen Aschoff (1918 – 1996) (1913 – 1998) Cold Spring Harbor Symposium on Quantitative Biology, Vol. XXV Biological Clocks The Circadian Circuit Environmental Output Inputs Rhythms Light Hormonal Cycles Temperature Rest/Wake Social Activity Feeding Central Pacemaker What would a circadian pacemaker look like? The Circadian Circuit Environmental Output Inputs Rhythms Light Hormonal Cycles Temperature -

Pittendrigh (1918-1996): in Fond Memory
ARTICLE-IH-A-BOX Pittendrigh (1918-1996): In Fond Memory The scientific study of biological rhythms (chronobiology) has really had a many faceted and colourful past in Europe and drew the attention of brilliant naturalists in the 18th, 19th and 20th centuries and is turning out to be one of the most interdiscipli nary branches of modem biology in the 21st century. The conceptual foundations of chronobiology in the 20th century were laid by Jurgen Aschoff, Erwin Blinning and Colin Pittendrigh. Their deep insight and contributions in the second half of that century account for the high degree of scientific elegance and experimental rigour of the subject. In fact, it was the symposium on Biological Clocks convened by Pittendrigh in the Cold Spring Harbor in New York in 1960, with Bunning as the chairperson and the seminal papers of Aschoff and Pittendrigh that ushered in an era of intensified experimental activity. Bunning's address was a definitive version of the history of chronobiology. By 1997, researchers began talking ofa 'party time' for chronobiology since it marked the twenty-fifth anniversary of the discovery of the supra chiasmatic nucleus (SeN) in the brain of mammals, which is the pacemaker of circadian organization, and the identification and cloning of the first circadian clock gene in a mammal. The journal Science, in a December 1998 issue, ranked some of the findings in the field of biological clocks as the first runner-up breakthrough-of-the-year. By 2003 one of our fraternity could write: "The successes achieved by chronobiologists over the past decade have been the envy of the scientific community. -

Clock Mutants of Drosophila Melanogaster (Eclosion/Circadian/Rhythms/X Chromosome) RONALD J
Proc. Nat. Acad. Sci. USA Vol. 68, No. 9, pp. 2112-2116, September 1971 Clock Mutants of Drosophila melanogaster (eclosion/circadian/rhythms/X chromosome) RONALD J. KONOPKA AND SEYMOUR BENZER Division of Biology, California Institute of Technology, Pasadena, Calif. 91109 Contributed by Seymour Benzer, July 2, 1971 ABSTRACT Three mutants have been isolated in dark period. In a few bottles, males emerged in approximately which the normal 24-hour rhythm is drastically changed. equal numbers during day and night. Each mutant candidate One mutant is arrhythmic; another has a period of 19 hr; a third has a period of 28 hr. Both the eclosion rhythm of a was examined in more detail by raising pupae in LD 12:12, population and the locomotor activity of individual flies then monitoring the adult eclosion rhythm in constant are affected. All these mutations appear to involve the darkness. From a total of about 2000 F1 males, three rhythm same functional gene on the X chromosome. mutants were obtained. Rhythmic variations in behavior are displayed by many Determination of eclosion and locomotor organisms, ranging from single cells to man (1). When the activity rhythms rhythm persists under constant conditions, and has a period Eclosion rhythms, free-running in constant darkness, were of around one day, depending little on temperature, the determined with automatic "bang boxes" (20), generously rhythm is called circadian (2). Many experiments have at- loaned by Dr. Colin Pittendrigh. Several hundred pupae, tempted to probe the mechanism (3), but the nature of the raised in LD 12:12, were transferred to the apparatus at the underlying oscillation remains unknown (4). -

Searches for Life and Intelligence Beyond Earth
Technologies of Perception: Searches for Life and Intelligence Beyond Earth by Claire Isabel Webb Bachelor of Arts, cum laude Vassar College, 2010 Submitted to the Program in Science, Technology and Society in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy in History, Anthropology, and Science, Technology and Society at the Massachusetts Institute of Technology September 2020 © 2020 Claire Isabel Webb. All Rights Reserved. The author hereby grants to MIT permission to reproduce and distribute publicly paper and electronic copies of this thesis document in whole or in part in any medium now known or hereafter created. Signature of Author: _____________________________________________________________ History, Anthropology, and Science, Technology and Society August 24, 2020 Certified by: ___________________________________________________________________ David Kaiser Germeshausen Professor of the History of Science (STS) Professor of Physics Thesis Supervisor Certified by: ___________________________________________________________________ Stefan Helmreich Elting E. Morison Professor of Anthropology Thesis Committee Member Certified by: ___________________________________________________________________ Sally Haslanger Ford Professor of Philosophy and Women’s and Gender Studies Thesis Committee Member Accepted by: ___________________________________________________________________ Graham Jones Associate Professor of Anthropology Director of Graduate Studies, History, Anthropology, and STS Accepted by: ___________________________________________________________________ -

Intrinsic and Extrinsic Regulation of Pineal Melatonin Rhythms
University of Kentucky UKnowledge Theses and Dissertations--Biology Biology 2016 INTRINSIC AND EXTRINSIC REGULATION OF PINEAL MELATONIN RHYTHMS Ye Li University of Kentucky, [email protected] Digital Object Identifier: http://dx.doi.org/10.13023/ETD.2016.108 Right click to open a feedback form in a new tab to let us know how this document benefits ou.y Recommended Citation Li, Ye, "INTRINSIC AND EXTRINSIC REGULATION OF PINEAL MELATONIN RHYTHMS" (2016). Theses and Dissertations--Biology. 34. https://uknowledge.uky.edu/biology_etds/34 This Doctoral Dissertation is brought to you for free and open access by the Biology at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Biology by an authorized administrator of UKnowledge. For more information, please contact [email protected]. STUDENT AGREEMENT: I represent that my thesis or dissertation and abstract are my original work. Proper attribution has been given to all outside sources. I understand that I am solely responsible for obtaining any needed copyright permissions. I have obtained needed written permission statement(s) from the owner(s) of each third-party copyrighted matter to be included in my work, allowing electronic distribution (if such use is not permitted by the fair use doctrine) which will be submitted to UKnowledge as Additional File. I hereby grant to The University of Kentucky and its agents the irrevocable, non-exclusive, and royalty-free license to archive and make accessible my work in whole or in part in all forms of media, now or hereafter known. I agree that the document mentioned above may be made available immediately for worldwide access unless an embargo applies. -

Journal of Circadian Rhythms Biomed Central
Journal of Circadian Rhythms BioMed Central Review Open Access Transdisciplinary unifying implications of circadian findings in the 1950s Franz Halberg*1, Germaine Cornélissen1, George Katinas1, Elena V Syutkina2, Robert B Sothern1, Rina Zaslavskaya3, Francine Halberg1, Yoshihiko Watanabe4, Othild Schwartzkopff1, Kuniaki Otsuka5, Roberto Tarquini6, Perfetto Frederico6 and Jarmila Siggelova7 Address: 1Halberg Chronobiology Center, University of Minnesota, Minneapolis, MN, USA, 2Institute of Pediatrics, Scientific Center for Children's Health, Academy of Medical Sciences, Moscow, Russia, 3Department of Cardiology, Hospital #60, Moscow, Russia, 4Tokyo Women's Medical University, Daini Hospital, Tokyo, Japan, 5Tokyo Women Medical University, School of Medicine, Daini Hospital, Division of Neurocardiology and Chronoecology, Nishiogu 2-1-10, Arakawa-ku, Tokyo 116-856, Japan, 6Department of Internal Medicine, University of Florence, Italy and 7Clinic of Functional Diagnostics and Rehabilitation, St. Anna Faculty Hospital and Masaryk University of Brno, Pekaská 53, 656 91, Brno, Czech Republic Email: Franz Halberg* - [email protected]; Germaine Cornélissen - [email protected]; George Katinas - [email protected]; Elena V Syutkina - [email protected]; Robert B Sothern - [email protected]; Rina Zaslavskaya - [email protected]; Francine Halberg - [email protected]; Yoshihiko Watanabe - [email protected]; Othild Schwartzkopff - [email protected]; Kuniaki Otsuka - [email protected]_net.ne.jp; Roberto Tarquini - [email protected]; Perfetto Frederico - [email protected]; Jarmila Siggelova - [email protected] * Corresponding author Published: 29 October 2003 Received: 24 September 2003 Accepted: 29 October 2003 Journal of Circadian Rhythms 2003, 1:2 This article is available from: http://www.JCircadianRhythms.com/content/1/1/2 © 2003 Halberg et al; licensee BioMed Central Ltd. -
Colin Pittendrigh in His Final Years at Princeton University
Colin Pittendrigh in his final years at Princeton University The manuscript associated with these remarks is an abbreviated version of my Ph.D. dissertation, earned in 1969 under the guidance of Colin Pittendrigh (Pitt) at Princeton University. The manuscript was not published, or more accurately was rejected, by Science magazine in 1976. I chose the subject because I was interested in the ecology of flies of the genus Drosophila, and I wanted to generate evidence that natural environmental conditions are a source of selection for the well known circadian eclosion rhythm of these animals. All species of Drosophila tested have a circadian eclosion rhythm that causes adult flies to emerge from the pupal case only in the first few hours after dawn. I selected one species of Drosophila native to the western U.S., and a second species native to the eastern U.S, and exposed both to natural and extreme saturation deficit regimes. Saturation deficit is a measure of the evaporative power of the atmosphere at a particular temperature and relative humidity. I wanted to find out if Drosophila in nature more successfully eclose in the early morning because that is the daylight time when the saturation deficit is usually at a minimum. I found this to be true for both species when exposed to their natural environment. In the laboratory, I found a positive relationship between eclosion success and low saturation deficits. Pitt steadily encouraged me in this work, reviewed the manuscript and made helpful suggestions before I submitted it for publication. I arrived at Princeton in the fall of 1965, eager to meet my new scientific mentor, Colin Pittendrigh. -

Historical Studies in the Societal Impact of Spaceflight
Historical Studies in the Societal Impact of Spaceflight Steven J. Dick Editor Historical Studies in the Societal Impact of Spaceflight Historical Studies in the Societal Impact of Spaceflight Steven J. Dick Editor National Aeronautics and Space Administration Office of Communications NASA History Program Office Washington, DC 2015 NASA SP-2015-4803 Library of Congress Cataloging-in-Publication Data Historical studies in the societal impact of spaceflight / Steven J. Dick, editor. p. cm. — (Societal impact series ; v. 3) Includes bibliographical references and index. “NASA SP-2015-4803.” 1. Astronautics—Technology transfer—History. 2. Astronautics—Public opinion—History. 3. Astronautics—Social aspects—History. 4. United States. National Aeronautics and Space Administration. I. Dick, Steven J. TL865.H58 2010 338.973’06--dc22 2009030014 This publication is available as a free download at http://www.nasa.gov/ebooks. v Contents Introduction ..................................vii PART I OPINION Chapter 1. The Impact of Space Exploration on Public Opinions, Attitudes, and Beliefs William Sims Bainbridge .................................1 PART II SPINOFF? Chapter 2. Societal Impact of NASA on Medical Technology William Sims Bainbridge ................................77 Chapter 3. NASA’s Role in the Manufacture of Integrated Circuits Andrew J. Butrica ....................................149 Chapter 4. NASA’s Role in the Development of MEMS (Microelectromechanical Systems) Andrew J. Butrica ....................................251 PART III THE WORLD AT LARGE Chapter 5. Powering Space Exploration: U.S. Space Nuclear Power, Public Perceptions, and Outer Planetary Probes Roger D. Launius ....................................331 Chapter 6. NASA and the Environment: An Evolving Relationship W. Henry Lambright ..................................383 Chapter 7. Societal Impacts of Applications Satellites David J. Whalen .....................................427 vi Historical Studies in the Societal Impact of Spaceflight Chapter 8. -

The Program in Human Biology at Stanford University the First 30 Years
The Program in Human Biology at Stanford University The first 30 years ‒ HUMAN BIOLOGY CORE COURSES AND FACULTY COORDINATORS Sciences: Donald Kennedy 3B The Social Sciences:Arthur Wolf, John Gurley 1969/70 4A The Biological Sciences: Merton Bernfield 1 Man and Nature: Colin Pittendrigh 4B. The Social Sciences: Herant Katchadourian 1970/71 1977/78 1 Man and Nature: Colin Pittendrigh 2A Basic Concepts in Biology: Merton Bernfield, Richard Goldsby 2A Cells, Organisms, and Societies: Colin Pittendrigh 2B Basic Concepts in the Social Sciences: Arthur Wolf, Sandy Dornbusch 2B Behavior As Adaptation: David Hamburg, Alberta Siegel, Herant 3A The Biological Sciences: Jeffrey Wine, Merton Bernfield Katchadourian, Sandy Dornbusch 3B The Social Sciences: Albert Hastorf, Sandy Dornbusch, Shirley Feldman 3A Man As an Organism: Norman Kretchmer 4A The Biological Sciences: William Durham 3B The Transformation of Human Society: Sandy Dornbusch, John Gurley, 4B The Social Sciences: Arthur Wolf David Hamburg, Alberta Siegel, Herant Katchadourian 1978/79 4A Biology of Populations: Paul Ehrlich 2A & 2B Human Evolution: Culture and the Cell: Merton Bernfield (2A); 4B Topics in Sociobiology: Sandy Dornbusch, David Hamburg, Herant Arthur Wolf (2B) Katchadourian, Alberta Siegel 3A & 3B The Human Organism: Principles of Social-Psychological and 1971/72 Physiological Regulation and Integration: Merton Bernfield (3A); 1 Man and Nature: Colin Pittendrigh Sandy Dornbusch, Shirley Feldman (3B) 2A Cells, Organisms, and Societies: Donald Kennedy 4A & 4B Populations and -

CIRCADIAN PHYSIOLOGY Third Edition This Page Intentionally Left Blank CIRCADIAN PHYSIOLOGY Third Edition
CIRCADIAN PHYSIOLOGY Third Edition This page intentionally left blank CIRCADIAN PHYSIOLOGY Third Edition Roberto Refinetti, PhD Boise State University Idaho, USA CRC Press Taylor & Francis Group 6000 Broken Sound Parkway NW, Suite 300 Boca Raton, FL 33487-2742 © 2016 by Taylor & Francis Group, LLC CRC Press is an imprint of Taylor & Francis Group, an Informa business No claim to original U.S. Government works Version Date: 20160202 International Standard Book Number-13: 978-1-4665-1498-0 (eBook - PDF) This book contains information obtained from authentic and highly regarded sources. Reasonable efforts have been made to publish reliable data and information, but the author and publisher cannot assume responsibility for the validity of all materials or the consequences of their use. The authors and publishers have attempted to trace the copyright holders of all material reproduced in this publication and apologize to copyright holders if permission to publish in this form has not been obtained. If any copyright material has not been acknowledged please write and let us know so we may rectify in any future reprint. Except as permitted under U.S. Copyright Law, no part of this book may be reprinted, reproduced, transmitted, or utilized in any form by any electronic, mechanical, or other means, now known or hereafter invented, including photocopying, microfilming, and recording, or in any information storage or retrieval system, without written permission from the publishers. For permission to photocopy or use material electronically from this work, please access www.copyright.com (http://www.copyright.com/) or contact the Copyright Clearance Center, Inc. (CCC), 222 Rosewood Drive, Danvers, MA 01923, 978-750-8400.