Botany Without Bias
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Ecological Correlates of Ghost Lineages in Ruminants
Paleobiology, 38(1), 2012, pp. 101–111 Ecological correlates of ghost lineages in ruminants Juan L. Cantalapiedra, Manuel Herna´ndez Ferna´ndez, Gema M. Alcalde, Beatriz Azanza, Daniel DeMiguel, and Jorge Morales Abstract.—Integration between phylogenetic systematics and paleontological data has proved to be an effective method for identifying periods that lack fossil evidence in the evolutionary history of clades. In this study we aim to analyze whether there is any correlation between various ecomorphological variables and the duration of these underrepresented portions of lineages, which we call ghost lineages for simplicity, in ruminants. Analyses within phylogenetic (Generalized Estimating Equations) and non-phylogenetic (ANOVAs and Pearson correlations) frameworks were performed on the whole phylogeny of this suborder of Cetartiodactyla (Mammalia). This is the first time ghost lineages are focused in this way. To test the robustness of our data, we compared the magnitude of ghost lineages among different continents and among phylogenies pruned at different ages (4, 8, 12, 16, and 20 Ma). Differences in mean ghost lineage were not significantly related to either geographic or temporal factors. Our results indicate that the proportion of the known fossil record in ruminants appears to be influenced by the preservation potential of the bone remains in different environments. Furthermore, large geographical ranges of species increase the likelihood of preservation. Juan L. Cantalapiedra, Gema Alcalde, and Jorge Morales. Departamento de Paleobiologı´a, Museo Nacional de Ciencias Naturales, UCM-CSIC, Pinar 25, 28006 Madrid, Spain. E-mail: [email protected] Manuel Herna´ndez Ferna´ndez. Departamento de Paleontologı´a, Facultad de Ciencias Geolo´gicas, Universidad Complutense de Madrid y Departamento de Cambio Medioambiental, Instituto de Geociencias, Consejo Superior de Investigaciones Cientı´ficas, Jose´ Antonio Novais 2, 28040 Madrid, Spain Daniel DeMiguel. -

Lecture Notes: the Mathematics of Phylogenetics
Lecture Notes: The Mathematics of Phylogenetics Elizabeth S. Allman, John A. Rhodes IAS/Park City Mathematics Institute June-July, 2005 University of Alaska Fairbanks Spring 2009, 2012, 2016 c 2005, Elizabeth S. Allman and John A. Rhodes ii Contents 1 Sequences and Molecular Evolution 3 1.1 DNA structure . .4 1.2 Mutations . .5 1.3 Aligned Orthologous Sequences . .7 2 Combinatorics of Trees I 9 2.1 Graphs and Trees . .9 2.2 Counting Binary Trees . 14 2.3 Metric Trees . 15 2.4 Ultrametric Trees and Molecular Clocks . 17 2.5 Rooting Trees with Outgroups . 18 2.6 Newick Notation . 19 2.7 Exercises . 20 3 Parsimony 25 3.1 The Parsimony Criterion . 25 3.2 The Fitch-Hartigan Algorithm . 28 3.3 Informative Characters . 33 3.4 Complexity . 35 3.5 Weighted Parsimony . 36 3.6 Recovering Minimal Extensions . 38 3.7 Further Issues . 39 3.8 Exercises . 40 4 Combinatorics of Trees II 45 4.1 Splits and Clades . 45 4.2 Refinements and Consensus Trees . 49 4.3 Quartets . 52 4.4 Supertrees . 53 4.5 Final Comments . 54 4.6 Exercises . 55 iii iv CONTENTS 5 Distance Methods 57 5.1 Dissimilarity Measures . 57 5.2 An Algorithmic Construction: UPGMA . 60 5.3 Unequal Branch Lengths . 62 5.4 The Four-point Condition . 66 5.5 The Neighbor Joining Algorithm . 70 5.6 Additional Comments . 72 5.7 Exercises . 73 6 Probabilistic Models of DNA Mutation 81 6.1 A first example . 81 6.2 Markov Models on Trees . 87 6.3 Jukes-Cantor and Kimura Models . -

Phylogeny of a Rapidly Evolving Clade: the Cichlid Fishes of Lake Malawi
Proc. Natl. Acad. Sci. USA Vol. 96, pp. 5107–5110, April 1999 Evolution Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa (adaptive radiationysexual selectionyspeciationyamplified fragment length polymorphismylineage sorting) R. C. ALBERTSON,J.A.MARKERT,P.D.DANLEY, AND T. D. KOCHER† Department of Zoology and Program in Genetics, University of New Hampshire, Durham, NH 03824 Communicated by John C. Avise, University of Georgia, Athens, GA, March 12, 1999 (received for review December 17, 1998) ABSTRACT Lake Malawi contains a flock of >500 spe- sponsible for speciation, then we expect that sister taxa will cies of cichlid fish that have evolved from a common ancestor frequently differ in color pattern but not morphology. within the last million years. The rapid diversification of this Most attempts to determine the relationships among cichlid group has been attributed to morphological adaptation and to species have used morphological characters, which may be sexual selection, but the relative timing and importance of prone to convergence (8). Molecular sequences normally these mechanisms is not known. A phylogeny of the group provide the independent estimate of phylogeny needed to infer would help identify the role each mechanism has played in the evolutionary mechanisms. The Lake Malawi cichlids, however, evolution of the flock. Previous attempts to reconstruct the are speciating faster than alleles can become fixed within a relationships among these taxa using molecular methods have species (9, 10). The coalescence of mtDNA haplotypes found been frustrated by the persistence of ancestral polymorphisms within populations predates the origin of many species (11). In within species. -

Major Clades of Agaricales: a Multilocus Phylogenetic Overview
Mycologia, 98(6), 2006, pp. 982–995. # 2006 by The Mycological Society of America, Lawrence, KS 66044-8897 Major clades of Agaricales: a multilocus phylogenetic overview P. Brandon Matheny1 Duur K. Aanen Judd M. Curtis Laboratory of Genetics, Arboretumlaan 4, 6703 BD, Biology Department, Clark University, 950 Main Street, Wageningen, The Netherlands Worcester, Massachusetts, 01610 Matthew DeNitis Vale´rie Hofstetter 127 Harrington Way, Worcester, Massachusetts 01604 Department of Biology, Box 90338, Duke University, Durham, North Carolina 27708 Graciela M. Daniele Instituto Multidisciplinario de Biologı´a Vegetal, M. Catherine Aime CONICET-Universidad Nacional de Co´rdoba, Casilla USDA-ARS, Systematic Botany and Mycology de Correo 495, 5000 Co´rdoba, Argentina Laboratory, Room 304, Building 011A, 10300 Baltimore Avenue, Beltsville, Maryland 20705-2350 Dennis E. Desjardin Department of Biology, San Francisco State University, Jean-Marc Moncalvo San Francisco, California 94132 Centre for Biodiversity and Conservation Biology, Royal Ontario Museum and Department of Botany, University Bradley R. Kropp of Toronto, Toronto, Ontario, M5S 2C6 Canada Department of Biology, Utah State University, Logan, Utah 84322 Zai-Wei Ge Zhu-Liang Yang Lorelei L. Norvell Kunming Institute of Botany, Chinese Academy of Pacific Northwest Mycology Service, 6720 NW Skyline Sciences, Kunming 650204, P.R. China Boulevard, Portland, Oregon 97229-1309 Jason C. Slot Andrew Parker Biology Department, Clark University, 950 Main Street, 127 Raven Way, Metaline Falls, Washington 99153- Worcester, Massachusetts, 01609 9720 Joseph F. Ammirati Else C. Vellinga University of Washington, Biology Department, Box Department of Plant and Microbial Biology, 111 355325, Seattle, Washington 98195 Koshland Hall, University of California, Berkeley, California 94720-3102 Timothy J. -

Phylogenetic Comparative Methods: a User's Guide for Paleontologists
Phylogenetic Comparative Methods: A User’s Guide for Paleontologists Laura C. Soul - Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA David F. Wright - Division of Paleontology, American Museum of Natural History, Central Park West at 79th Street, New York, New York 10024, USA and Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington, DC, USA Abstract. Recent advances in statistical approaches called Phylogenetic Comparative Methods (PCMs) have provided paleontologists with a powerful set of analytical tools for investigating evolutionary tempo and mode in fossil lineages. However, attempts to integrate PCMs with fossil data often present workers with practical challenges or unfamiliar literature. In this paper, we present guides to the theory behind, and application of, PCMs with fossil taxa. Based on an empirical dataset of Paleozoic crinoids, we present example analyses to illustrate common applications of PCMs to fossil data, including investigating patterns of correlated trait evolution, and macroevolutionary models of morphological change. We emphasize the importance of accounting for sources of uncertainty, and discuss how to evaluate model fit and adequacy. Finally, we discuss several promising methods for modelling heterogenous evolutionary dynamics with fossil phylogenies. Integrating phylogeny-based approaches with the fossil record provides a rigorous, quantitative perspective to understanding key patterns in the history of life. 1. Introduction A fundamental prediction of biological evolution is that a species will most commonly share many characteristics with lineages from which it has recently diverged, and fewer characteristics with lineages from which it diverged further in the past. This principle, which results from descent with modification, is one of the most basic in biology (Darwin 1859). -

Complete Plastome Sequences Of
Karol et al. BMC Evolutionary Biology 2010, 10:321 http://www.biomedcentral.com/1471-2148/10/321 RESEARCH ARTICLE Open Access Complete plastome sequences of Equisetum arvense and Isoetes flaccida: implications for phylogeny and plastid genome evolution of early land plant lineages Kenneth G Karol1*, Kathiravetpillai Arumuganathan2, Jeffrey L Boore3,4, Aaron M Duffy5, Karin DE Everett6, John D Hall1, S Kellon Hansen5, Jennifer V Kuehl7, Dina F Mandoli6,8, Brent D Mishler9, Richard G Olmstead6, Karen S Renzaglia10, Paul G Wolf5 Abstract Background: Despite considerable progress in our understanding of land plant phylogeny, several nodes in the green tree of life remain poorly resolved. Furthermore, the bulk of currently available data come from only a subset of major land plant clades. Here we examine early land plant evolution using complete plastome sequences including two previously unexamined and phylogenetically critical lineages. To better understand the evolution of land plants and their plastomes, we examined aligned nucleotide sequences, indels, gene and nucleotide composition, inversions, and gene order at the boundaries of the inverted repeats. Results: We present the plastome sequences of Equisetum arvense, a horsetail, and of Isoetes flaccida,a heterosporous lycophyte. Phylogenetic analysis of aligned nucleotides from 49 plastome genes from 43 taxa supported monophyly for the following clades: embryophytes (land plants), lycophytes, monilophytes (leptosporangiate ferns + Angiopteris evecta + Psilotum nudum + Equisetum arvense), and seed plants. Resolution among the four monilophyte lineages remained moderate, although nucleotide analyses suggested that P. nudum and E. arvense form a clade sister to A. evecta + leptosporangiate ferns. Results from phylogenetic analyses of nucleotides were consistent with the distribution of plastome gene rearrangements and with analysis of sequence gaps resulting from insertions and deletions (indels). -

Early Tetrapod Relationships Revisited
Biol. Rev. (2003), 78, pp. 251–345. f Cambridge Philosophical Society 251 DOI: 10.1017/S1464793102006103 Printed in the United Kingdom Early tetrapod relationships revisited MARCELLO RUTA1*, MICHAEL I. COATES1 and DONALD L. J. QUICKE2 1 The Department of Organismal Biology and Anatomy, The University of Chicago, 1027 East 57th Street, Chicago, IL 60637-1508, USA ([email protected]; [email protected]) 2 Department of Biology, Imperial College at Silwood Park, Ascot, Berkshire SL57PY, UK and Department of Entomology, The Natural History Museum, Cromwell Road, London SW75BD, UK ([email protected]) (Received 29 November 2001; revised 28 August 2002; accepted 2 September 2002) ABSTRACT In an attempt to investigate differences between the most widely discussed hypotheses of early tetrapod relation- ships, we assembled a new data matrix including 90 taxa coded for 319 cranial and postcranial characters. We have incorporated, where possible, original observations of numerous taxa spread throughout the major tetrapod clades. A stem-based (total-group) definition of Tetrapoda is preferred over apomorphy- and node-based (crown-group) definitions. This definition is operational, since it is based on a formal character analysis. A PAUP* search using a recently implemented version of the parsimony ratchet method yields 64 shortest trees. Differ- ences between these trees concern: (1) the internal relationships of aı¨stopods, the three selected species of which form a trichotomy; (2) the internal relationships of embolomeres, with Archeria -

Phylogenetics: Recovering Evolutionary History COMP 571 Luay Nakhleh, Rice University
1 Phylogenetics: Recovering Evolutionary History COMP 571 Luay Nakhleh, Rice University 2 The Structure and Interpretation of Phylogenetic Trees unrooted, binary species tree rooted, binary species tree speciation (direction of descent) Flow of time ๏ six extant taxa or operational taxonomic units (OTUs) 3 The Structure and Interpretation of Phylogenetic Trees Phylogenetics-RecoveringEvolutionaryHistory - March 3, 2017 4 The Structure and Interpretation of Phylogenetic Trees In a binary tree on n taxa, how may nodes, branches, internal nodes and internal branches are there? How many unrooted binary trees on n taxa are there? How many rooted binary trees on n taxa are there? ๏ six extant taxa or operational taxonomic units (OTUs) 5 The Structure and Interpretation of Phylogenetic Trees polytomy Non-binary Multifuracting Partially resolved Polytomous ๏ six extant taxa or operational taxonomic units (OTUs) 6 The Structure and Interpretation of Phylogenetic Trees A polytomy in a tree can be resolved (not necessarily fully) in many ways, thus producing trees with higher resolution (including binary trees) A binary tree can be turned into a partially resolved tree by contracting edges In how many ways can a polytomy of degree d be resolved? Compatibility between two trees guarantees that one can back and forth between the two trees by means of node refinement and edge contraction Phylogenetics-RecoveringEvolutionaryHistory - March 3, 2017 7 The Structure and Interpretation of Phylogenetic Trees branch lengths have Additive no meaning tree Additive tree ultrametric rooted at an tree outgroup (molecular clock) 8 The Structure and Interpretation of Phylogenetic Trees bipartition (split) AB|CDEF clade cluster 11 clades (4 nontrivial) 9 bipartitions (3 nontrivial) How many nontrivial clades are there in a binary tree on n taxa? How many nontrivial bipartitions are there in a binary tree on n taxa? How many possible nontrivial clusters of n taxa are there? 9 The Structure and Interpretation of Phylogenetic Trees Species vs. -

Brown Algae and 4) the Oomycetes (Water Molds)
Protista Classification Excavata The kingdom Protista (in the five kingdom system) contains mostly unicellular eukaryotes. This taxonomic grouping is polyphyletic and based only Alveolates on cellular structure and life styles not on any molecular evidence. Using molecular biology and detailed comparison of cell structure, scientists are now beginning to see evolutionary SAR Stramenopila history in the protists. The ongoing changes in the protest phylogeny are rapidly changing with each new piece of evidence. The following classification suggests 4 “supergroups” within the Rhizaria original Protista kingdom and the taxonomy is still being worked out. This lab is looking at one current hypothesis shown on the right. Some of the organisms are grouped together because Archaeplastida of very strong support and others are controversial. It is important to focus on the characteristics of each clade which explains why they are grouped together. This lab will only look at the groups that Amoebozoans were once included in the Protista kingdom and the other groups (higher plants, fungi, and animals) will be Unikonta examined in future labs. Opisthokonts Protista Classification Excavata Starting with the four “Supergroups”, we will divide the rest into different levels called clades. A Clade is defined as a group of Alveolates biological taxa (as species) that includes all descendants of one common ancestor. Too simplify this process, we have included a cladogram we will be using throughout the SAR Stramenopila course. We will divide or expand parts of the cladogram to emphasize evolutionary relationships. For the protists, we will divide Rhizaria the supergroups into smaller clades assigning them artificial numbers (clade1, clade2, clade3) to establish a grouping at a specific level. -
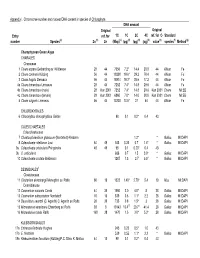
Green Appendix I
Appendix I. Chromsome number and nuclear DNA content in species of Chlorophyta DNA amount Original Original Entry ref. for 1C 1C 2C 4C ref. for C- Standard number Species(a) 2n (b) 2n (Mbp)(c) (pg)(d) (pg)(d) (pg)(d) value(e) species(f) Method(g) Charophycean Green Algae CHARALES Characeae 1 Chara aspera Detharding ex Willdenow 28 44 7056 7.2* 14.4 28.8 44 Allium Fe 2 Chara contraria Kützing 56 44 19208 19.6* 39.2 78.4 44 Allium Fe 3 Chara fragilis Desvaux 56 44 18914 19.3* 38.6 77.2 44 Allium Fe 4a Chara tomentosa Linnaeus 28 44 7252 7.4* 14.8 29.6 44 Allium Fe 4b Chara tomentosa (male) 28 Kun 2001 7252 7.4* 14.8 29.6 Kun 2001 Chara MI:EB 4c Chara tomentsoa (female) 28 Kun 2001 6860 7.0* 14.0 28.0 Kun 2001 Chara MI:EB 5 Chara vulgaris Linnaeus 56 44 13230 13.5* 27 54 44 Allium Fe CHLOROKYBALES 6 Chlorokybus atmosphyticus Geitler 98 0.1 0.2* 0.4 43 COLEOCHAETALES Coleochaetaceae 7 Chaetosphaeridium globosum (Nordstedt) Klebahn 1.2* * Gallus MI:DAPI 8 Coleochaete nitellarum Jost 84 49 343 0.35 0.7 1.4* * Gallus MI:DAPI 9a Coleochaete orbicularis Pringsheim 48 49 98 0.1 0.20* 0.4 43 9b C. orbicularis 686 0.7 1.5 3.0* * Gallus MI:DAPI 10 Coleochaete scutata Brébisson 1287 1.3 2.7 5.5* * Gallus MI:DAPI DESMIDIALES1 Closteriaceae 11 Closterium ehrenbergii Meneghini ex Ralfs 60 19 1323 1.40* 2.70* 5.4 19 Mus MI:DAPI Desmidiaceae 12 Cosmarium cucumis Corda 44 39 1960 2.0 4.0* 8 28 Gallus MI:DAPI 13 Cosmarium subcostatum Nordstedt 10 16 539 0.6 1.1* 2.2 28 Gallus MI:DAPI 14 Desmidium swartzii (C. -

Tempo and Mode in the Macroevolutionary Reconstruction of Darwinism STEPHEN JAY GOULD Museum of Comparative Zoology, Harvard University, Cambridge, MA 02138
Proc. Nadl. Acad. Sci. USA Vol. 91, pp. 6764-6771, July 1994 Colloquium Paper This paper was presented at a coloquium ented "Tempo and Mode in Evolution" organized by Walter M. Fitch and Francisco J. Ayala, held January 27-29, 1994, by the National Academy of Sciences, in Irvine, CA. Tempo and mode in the macroevolutionary reconstruction of Darwinism STEPHEN JAY GOULD Museum of Comparative Zoology, Harvard University, Cambridge, MA 02138 ABSTRACT Among the several central nings of Dar- But conceptual complexity is not reducible to a formula or winism, his version ofLyellian uniformitranism-the extrap- epigram (as we taxonomists of life's diversity should know olationist commitment to viewing causes ofsmall-scale, observ- better than most). Too much ink has been wasted in vain able change in modern populations as the complete source, by attempts to define the essence ofDarwin's ideas, or Darwin- smooth extension through geological time, of all magnitudes ism itself. Mayr (1) has correctly emphasized that many and sequences in evolution-has most contributed to the causal different, if related, Darwinisms exist, both in the thought of hegemony of microevolutlon and the assumption that paleon- the eponym himself, and in the subsequent history of evo- tology can document the contingent history of life but cannot lutionary biology-ranging from natural selection, to genea- act as a domain of novel evolutionary theory. G. G. Simpson logical connection of all living beings, to gradualism of tried to combat this view of paleontology as theoretically inert change. in his classic work, Tempo and Mode in Evolution (1944), with It would therefore be fatuous to claim that any one legit- a brilliant argument that the two subjects of his tide fall into a imate "essence" can be more basic or important than an- unue paleontological domain and that modes (processes and other. -

Multiple Origins of Life (Extinction/Bioclade/Precambrian/Evolution/Stochastic Processes) DAVID M
Proc. Natt Acad. Sci. USA Vol. 80; pp. 2981-2984, May 1983 Evolution Multiple origins of life (extinction/bioclade/Precambrian/evolution/stochastic processes) DAVID M. RAUP* AND JAMES W. VALENTINEt *Department of the Geophysical Sciences, University of Chicago, Chicago, Illinois 60637; and tDepartment of Geological Sciences, University of California, Santa Barbara, California 93106 Contributed by David M. Raup, February 22, 1983 ABSTRACT There is some indication that life may have orig- ing a full assortment of prebiotically synthesized organic build- inated readily under primitive earth conditions. If there were ing blocks for life and conditions appropriate for life origins, it multiple origins of life, the result could have been a polyphyletic is possible that rates of bioclade origins might compare well biota today. Using simple stochastic models for diversification and enough with rates of diversification. Indeed, there has been extinction,-we conclude: (i) the probability of survival of life is low speculation that life may have been polyphyletic (1, 4). How- unless there are multiple origins, and (ii) given survival of life and ever, there is strong evidence that all living forms are de- given as many as 10 independent origins of life, the odds are that scended from a single ancestor. Biochemical and organizational all but one would have gone extinct, yielding the monophyletic biota similarities and the "universality" of the genetic code indicate we have now. The fact of the survival of our particular form of this. Therefore, the question for this paper is: If there were life does not imply that it was unique or superior. multiple bioclades early in life history, what is the probability that only one would have survived? In other words, if life The formation of life de novo is generally viewed as unlikely or orig- impossible under present earth conditions.