Ecosystems Mario V
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Model for the Oceanic Mass Balance of Rhenium and Implications for the Extent of Proterozoic Ocean Anoxia
UC Riverside UC Riverside Previously Published Works Title A model for the oceanic mass balance of rhenium and implications for the extent of Proterozoic ocean anoxia Permalink https://escholarship.org/uc/item/1gh1h920 Journal GEOCHIMICA ET COSMOCHIMICA ACTA, 227 ISSN 0016-7037 Authors Sheen, Alex I Kendall, Brian Reinhard, Christopher T et al. Publication Date 2018-04-15 DOI 10.1016/j.gca.2018.01.036 Peer reviewed eScholarship.org Powered by the California Digital Library University of California Available online at www.sciencedirect.com ScienceDirect Geochimica et Cosmochimica Acta 227 (2018) 75–95 www.elsevier.com/locate/gca A model for the oceanic mass balance of rhenium and implications for the extent of Proterozoic ocean anoxia Alex I. Sheen a,b,⇑, Brian Kendall a, Christopher T. Reinhard c, Robert A. Creaser b, Timothy W. Lyons d, Andrey Bekker d, Simon W. Poulton e, Ariel D. Anbar f,g a Department of Earth and Environmental Sciences, University of Waterloo, 200 University Avenue West, Waterloo, Ontario N2L 3G1, Canada b Department of Earth and Atmospheric Sciences, University of Alberta, Edmonton, Alberta T6G 2E3, Canada c School of Earth and Atmospheric Sciences, Georgia Institute of Technology, Atlanta, GA 30332, USA d Department of Earth Sciences, University of California, Riverside, CA 92521, USA e School of Earth and Environment, University of Leeds, Leeds LS2 9JT, UK f School of Earth and Space Exploration, Arizona State University, Tempe, AZ 85287, USA g School of Molecular Sciences, Arizona State University, Tempe, AZ 85287, USA Received 13 July 2017; accepted in revised form 30 January 2018; available online 8 February 2018 Abstract Emerging geochemical evidence suggests that the atmosphere-ocean system underwent a significant decrease in O2 content following the Great Oxidation Event (GOE), leading to a mid-Proterozoic ocean (ca. -

Risk Analysis: Vessel Biofouling
Risk Analysis: Vessel Biofouling ISBN 978-0-478-37548-0 (print) ISBN 978-0-478-37549-7 (online) 15 February 2011 Risk Analysis: Vessel Biofouling 15 February 2011 Approved for general release Christine Reed Manager, Risk Analysis Ministry of Agriculture and Forestry Requests for further copies should be directed to: Publication Adviser MAF Information Bureau P O Box 2526 WELLINGTON Telephone: 0800 00 83 33 Facsimile: 04-894 0300 This publication is also available on the MAF website at http://www.biosecurity.govt.nz/regs/imports/ihs/risk © Crown Copyright - Ministry of Agriculture and Forestry i Contributors to this risk analysis 1. Primary author/s Dr Andrew Bell Senior Adviser MAF Biosecurity New Zealand Risk Analysis, Marine Wellington Simon Phillips Adviser MAF Biosecurity New Zealand Risk Analysis, Marine Wellington Dr Eugene Georgiades Senior Adviser MAF Biosecurity New Zealand Risk Analysis, Marine Wellington Dr Daniel Kluza Senior Adviser MAF Biosecurity New Zealand Risk Analysis, Marine Wellington 2. Secondary contributors Dr Christopher Denny Adviser MAF Biosecurity New Zealand Border Standards Wellington 3. External peer review John Lewis Principal Marine Consultant ES Link Services Pty Ltd Melbourne, Victoria, Australia Richard Piola Senior Scientist Cawthron Institute Nelson, New Zealand The draft risk analysis has also been internally reviewed by: Liz Jones (Border Standards); Justin McDonald (Post-Clearance); Melanie Newfield (Risk Analysis); Howard Pharo (Risk Analysis); Sandy Toy (Risk Analysis). The contribution of all the reviewers is gratefully acknowledged. ii Contents Page Executive summary 1 Definitions 7 1. Introduction 8 1.1. Background 8 1.2. Scope 13 1.3. References 14 2. Methodology 19 2.1. -

Cascading of Habitat Degradation: Oyster Reefs Invaded by Refugee Fishes Escaping Stress
Ecological Applications, 11(3), 2001, pp. 764±782 q 2001 by the Ecological Society of America CASCADING OF HABITAT DEGRADATION: OYSTER REEFS INVADED BY REFUGEE FISHES ESCAPING STRESS HUNTER S. LENIHAN,1 CHARLES H. PETERSON,2 JAMES E. BYERS,3 JONATHAN H. GRABOWSKI,2 GORDON W. T HAYER, AND DAVID R. COLBY NOAA-National Ocean Service, Center for Coastal Fisheries and Habitat Research, 101 Pivers Island Road, Beaufort, North Carolina 28516 USA Abstract. Mobile consumers have potential to cause a cascading of habitat degradation beyond the region that is directly stressed, by concentrating in refuges where they intensify biological interactions and can deplete prey resources. We tested this hypothesis on struc- turally complex, species-rich biogenic reefs created by the eastern oyster, Crassostrea virginica, in the Neuse River estuary, North Carolina, USA. We (1) sampled ®shes and invertebrates on natural and restored reefs and on sand bottom to compare ®sh utilization of these different habitats and to characterize the trophic relations among large reef-as- sociated ®shes and benthic invertebrates, and (2) tested whether bottom-water hypoxia and ®shery-caused degradation of reef habitat combine to induce mass emigration of ®sh that then modify community composition in refuges across an estuarine seascape. Experimen- tally restored oyster reefs of two heights (1 m tall ``degraded'' or 2 m tall ``natural'' reefs) were constructed at 3 and 6 m depths. We sampled hydrographic conditions within the estuary over the summer to monitor onset and duration of bottom-water hypoxia/anoxia, a disturbance resulting from density strati®cation and anthropogenic eutrophication. Reduc- tion of reef height caused by oyster dredging exposed the reefs located in deep water to hypoxia/anoxia for .2 wk, killing reef-associated invertebrate prey and forcing mobile ®shes into refuge habitats. -

Son of Perdition Free Download
SON OF PERDITION FREE DOWNLOAD Wendy Alec | 490 pages | 01 Apr 2010 | Warboys Publishing (Ireland) Limited | 9780956333001 | English | Dublin, Ireland Who Is the Son of Perdition? The unembodied spirits who supported Lucifer in the war in heaven and were cast out Moses and mortals who commit Son of Perdition unpardonable sin against the Holy Ghost will inherit the same condition as Lucifer and Cain, and thus are called "sons of Son of Perdition. Contrasting beliefs. Recently Popular Media x. Verse Only. BLB Searches. So who is he? The bible describes the behavior of one who worships Jesus Christ. The purposes of the Son of Perdition oppose those of Jesus. A verification email has been sent to the address you provided. Blue Letter Bible study tools make reading, Son of Perdition and studying the Bible easy and rewarding. Extinction event Holocene extinction Human extinction List of extinction events Genetic erosion Son of Perdition pollution. Christianity portal. Re-type Password. Even a believer who maintains the faith can become discouraged and lose the peace of the Holy Spirit when he is cut off from the body of Christ. Biblical texts. By proceeding, you consent to our cookie usage. Back Psalms 1. The Son of Perdition is the progeny of abuse and desolation. Sort Canonically. The Millennium. No Number. Advanced Options Exact Match. However, scripture declares that "the soul can never die" Son of Perdition and that in the Resurrection the spirit and the body are united "never to be divided" Alma ; cf. Individual instructors or editors may still require Son of Perdition use of URLs. -

View .Pdf of This Issue
The Hardy Orchid Society Newsletter No. 21 July 2001 The Hardy Orchid Society Committee is… President: Richard M Bateman Vice-Presidents: Paul Harcourt Davies and Norman Heywood Chairman: Richard Manuel, Wye View Cottage, Leys Hill, Ross-on-Wye, Herefordshire, HR9 5QU Secretary: Sarah Marks, 83 Ladysmith, East Gomeldon, Salisbury, Wilts, SP4 6LE Treasurer: Tony Beresford, Pound Farm, Wearne, Langport, Somerset, TA10 0QJ Membership Secretary: Nick Storer, 17 Orchard Close, Lymm, Cheshire, WA13 9HH Show Secretary: Doreen Webster, 25 Highfields Drive, Loughborough, Leics, LE11 3JS Newsletter Editor: Moira Tarrant, Bumby’s, Fox Rd., Mashbury, Chelmsford, CM1 4TJ Meetings Secretary: Colin Clay, 14 Cromwell Place, Lighthorne Heath, Leamington Spa, CV33 9TG Ordinary Member (publicity): Simon Tarrant, Bumby’s, Fox Rd., Mashbury, Chelmsford, CM1 4TJ Ordinary Member (Newsletter Dist.): Bill Temple, Primrose Cottage, Hanney Rd., Ste- venton, Oxon, OX13 6AP Ordinary Member (Seed & Fungus Bank): Ted Weeks, 74 Over Lane, Almondsbury, Bristol, BS32 4BT Co-opted Member (BOC Rep.): Richard Nicol, 1364 Evesham Rd., Astwood Bank, Red- ditch, Worcs, B96 6BD Contents P.3 From the New President, Richard Bateman P.5 Report of the 9th AGM of the Hardy Orchid Society P.7 Publicity Posters, Simon Tarrant P.8 HELP!, Richard Manuel P.8 HOS Plant Show 2001, Tony Hughes P.10 Does DNA reveal All about the Evolution of Terrestrial Orchids? Part 3, Richard Bateman P.14 Getting Started - the Basics of Hardy Orchid Cultivation, Alan Dash P.20 Chemical Warfare, Richard Manuel P.21 AGS Summer South Show - Silver Award, Carol Dash P.22 Seed and Fungus Bank, Ted Weeks Colour Insert between Pages 12 and 13 Cover illustration: Serapias lingua by Carol Dash HOS Newsletter 21, July 2001 From the New President of the HOS Richard Bateman I am of course delighted to accept the Presidency of the Hardy Orchid Society. -

Anatomia Associada Ao Comportamento Reprodutivo De
Jimena García Rodríguez Anatomia associada ao comportamento reprodutivo de Cubozoa Anatomy associated with the reproductive behavior of Cubozoa São Paulo 2015 Jimena García Rodríguez Anatomia associada ao comportamento reprodutivo de Cubozoa Anatomy associated with the reproductive behavior of Cubozoa Dissertação apresentada ao Instituto de Biociências da Universidade de São Paulo para obtenção de Título de Mestre em Ciências, na Área de Zoologia Orientador: Prof. Dr. Antonio Carlos Marques São Paulo 2015 García Rodríguez, Jimena Anatomia associada ao comportamento reprodutivo de Cubozoa 96 páginas Dissertação (Mestrado) - Instituto de Biociências da Universidade de São Paulo. Departamento de Zoologia. 1. Cubozoa; 2. Histologia; 3. Reprodução. I. Universidade de São Paulo. Instituto de Biociências. Departamento de Zoologia. Comissão Julgadora Prof(a) Dr(a) Prof(a) Dr(a) Prof. Dr. Antonio Carlos Marques A mis padres, hermana y en especial a mi abuelita “Caminante, son tus huellas el camino y nada más; Caminante, no hay camino, se hace camino al andar. Al andar se hace el camino, y al volver la vista atrás se ve la senda que nunca se ha de volver a pisar. Caminante no hay camino sino estelas en la mar” Antonio Machado, 1912 Agradecimentos Em primeiro lugar, eu gostaria de agradecer ao meu orientador Antonio Carlos Marques, Tim, pela confiança desde o primeiro dia, pela ajuda tanto pessoal como profissional durante os dois anos de mestrado, pelas discussões de cada tema tratado e estudado e pelas orientações que tornaram possível a elaboração deste trabalho. Agradeço também o apoio institucional do Instituto de Biociências e do Centro de Biologia Marinha da Universidade de São Paulo. -

Life Cycle Stages of the Benthic Palytoxin-Producing Dinoflagellate Ostreopsis Cf. Ovata (Dinophyceae)
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Digital.CSIC Life cycle stages of the benthic palytoxin-producing dinoflagellate Ostreopsis cf. ovata (Dinophyceae) Isabel Bravo1*, Magda Vila2, Silvia Casabianca3, Francisco Rodriguez1, Pilar Rial1, Pilar Riobó1, Antonella Penna3 1Unidad Asociada Fitoplancton Tóxico (CSIC-IEO), Instituto Español de Oceanografía (IEO). Subida a Radio Faro 50, 36390 Vigo, Spain 2Institut de Ciències del Mar (CSIC), Pg. Marítim de la Barceloneta 37-49, 08003 Barcelona, Spain. 3Department of Biomolecular Sciences, University of Urbino, V.le Trieste 296, 61100 Pesaro, Italy *E-mail address: [email protected] (I. Bravo) ABSTRACT The asexual and sexual reproduction of Ostreopsis cf. ovata was studied in the field and in cultures isolated from two locations in the Mediterranean Sea. Asexual division took place in the motile stage by the sharing of theca (desmoschisis). High cell-size variability and differences in division capability were detected in the cultures. Thecal analyses and nuclear division patterns allowed characterization of the different phases of dividing cells obtained during an in situ cell-cycle sampling performed off Llavaneres beach (Northeast Spain). During the 45-h cycle, binucleated cells accounted for 2.6% of the population. Division was initiated with the onset of dusk and reached a maximum 3–4 h before dawn. No dividing cells were detected after 09:00 AM. Sexuality occurred both in cultures and in natural populations of O. cf. ovata. Mating gamete pairs were the only sexual stages that could be distinguished from vegetative stages. The differences between these pairs and dividing cells are described herein. -

J. N. Adams – P. M. Brennan
J. N. ADAMS – P. M. BRENNAN THE TEXT AT LACTANTIUS, DE MORTIBUS PERSECUTORUM 44.2 AND SOME EPIGRAPHIC EVIDENCE FOR ITALIAN RECRUITS aus: Zeitschrift für Papyrologie und Epigraphik 84 (1990) 183–186 © Dr. Rudolf Habelt GmbH, Bonn 183 THE TEXT AT LACTANTIUS, DE MORTIBUS PERSECUTORUM 44.2 AND SOME EPIGRAPHIC EVIDENCE FOR ITALIAN RECRUITS At Lact. Mort. 44.2 the transmitted text (C = Codex Colbertinus, BN 2627) runs: 'plus uirium Maxentio erat, quod et patris sui exercitum receperat a Seuero et suum proprium de Mauris atque Italis nuper extraxerat'. Modern editors (Brandt-Laubmann, Moreau, Creed) have accepted Heumann's conjecture G(a)etulis for Italis without discussion.1 But is the change necessary? Much depends on the interpretation of extraxerat. Creed translates… '... his own (army), which he had recently brought over from the Mauri and the Gaetuli' (our italics). In his note (p.118) he finds here an allusion to 'the rebellion against Maxentius of L. Domitius Alexander, vicarius of Africa, which probably began in 308 and was probably crushed by the end of 309'. On this view the troops will have been 'brought over' from Africa after the rebellion was put down. Creed's translation is along the same lines as that of Moreau… I, p.126 'il venait de faire revenir la sienne propre du pays des Maures et des Gétules'.2 In their eagerness to find a reference to L. Domitius Alexander, Moreau and Creed have mistranslated extraxerat. Neither editor attempts to explain how a verb basically meaning 'drag out of' could acquire the sense 'bring over, cause to return'. -

Phylogenetics of Tribe Orchideae (Orchidaceae: Orchidoideae)
Annals of Botany 110: 71–90, 2012 doi:10.1093/aob/mcs083, available online at www.aob.oxfordjournals.org Phylogenetics of tribe Orchideae (Orchidaceae: Orchidoideae) based on combined DNA matrices: inferences regarding timing of diversification and evolution of pollination syndromes Luis A. Inda1,*, Manuel Pimentel2 and Mark W. Chase3 1Escuela Polite´cnica Superior de Huesca, Universidad de Zaragoza, carretera de Cuarte sn. 22071 Huesca, Spain, 2Facultade de Ciencias, Universidade da Corun˜a, Campus da Zapateira sn. 15071 A Corun˜a, Spain and 3Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK * For correspondence. E-mail [email protected] Received: 3 November 2011 Returned for revision: 9 December 2011 Accepted: 1 March 2012 Published electronically: 25 April 2012 † Background and aims Tribe Orchideae (Orchidaceae: Orchidoideae) comprises around 62 mostly terrestrial genera, which are well represented in the Northern Temperate Zone and less frequently in tropical areas of both the Old and New Worlds. Phylogenetic relationships within this tribe have been studied previously using only nuclear ribosomal DNA (nuclear ribosomal internal transcribed spacer, nrITS). However, different parts of the phylogenetic tree in these analyses were weakly supported, and integrating information from different plant genomes is clearly necessary in orchids, where reticulate evolution events are putatively common. The aims of this study were to: (1) obtain a well-supported and dated phylogenetic hypothesis for tribe Orchideae, (ii) assess appropriateness of recent nomenclatural changes in this tribe in the last decade, (3) detect possible examples of reticulate evolution and (4) analyse in a temporal context evolutionary trends for subtribe Orchidinae with special emphasis on pollination systems. -

Marine Mammals and Sea Turtles of the Mediterranean and Black Seas
Marine mammals and sea turtles of the Mediterranean and Black Seas MEDITERRANEAN AND BLACK SEA BASINS Main seas, straits and gulfs in the Mediterranean and Black Sea basins, together with locations mentioned in the text for the distribution of marine mammals and sea turtles Ukraine Russia SEA OF AZOV Kerch Strait Crimea Romania Georgia Slovenia France Croatia BLACK SEA Bosnia & Herzegovina Bulgaria Monaco Bosphorus LIGURIAN SEA Montenegro Strait Pelagos Sanctuary Gulf of Italy Lion ADRIATIC SEA Albania Corsica Drini Bay Spain Dardanelles Strait Greece BALEARIC SEA Turkey Sardinia Algerian- TYRRHENIAN SEA AEGEAN SEA Balearic Islands Provençal IONIAN SEA Syria Basin Strait of Sicily Cyprus Strait of Sicily Gibraltar ALBORAN SEA Hellenic Trench Lebanon Tunisia Malta LEVANTINE SEA Israel Algeria West Morocco Bank Tunisian Plateau/Gulf of SirteMEDITERRANEAN SEA Gaza Strip Jordan Suez Canal Egypt Gulf of Sirte Libya RED SEA Marine mammals and sea turtles of the Mediterranean and Black Seas Compiled by María del Mar Otero and Michela Conigliaro The designation of geographical entities in this book, and the presentation of the material, do not imply the expression of any opinion whatsoever on the part of IUCN concerning the legal status of any country, territory, or area, or of its authorities, or concerning the delimitation of its frontiers or boundaries. The views expressed in this publication do not necessarily reflect those of IUCN. Published by Compiled by María del Mar Otero IUCN Centre for Mediterranean Cooperation, Spain © IUCN, Gland, Switzerland, and Malaga, Spain Michela Conigliaro IUCN Centre for Mediterranean Cooperation, Spain Copyright © 2012 International Union for Conservation of Nature and Natural Resources With the support of Catherine Numa IUCN Centre for Mediterranean Cooperation, Spain Annabelle Cuttelod IUCN Species Programme, United Kingdom Reproduction of this publication for educational or other non-commercial purposes is authorized without prior written permission from the copyright holder provided the sources are fully acknowledged. -
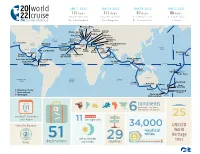
World Cruise - 2022 Use the Down Arrow from a Form Field
This document contains both information and form fields. To read information, World Cruise - 2022 use the Down Arrow from a form field. 20 world JAN 5, 2022 JAN 19, 2022 JAN 19, 2022 JAN 5, 2022 111 days 111 days 97 days 88 days 22 cruise roundtrip from roundtrip from Los Angeles to Ft. Lauderdale Ft. Lauderdale Los Angeles Ft. Lauderdale to Rome Florence/Pisa (Livorno) Genoa Rome (Civitavecchia) Catania Monte Carlo (Sicily) MONACO ITALY Naples Marseille Mykonos FRANCE GREECE Kusadasi PORTUGAL Atlantic Barcelona Heraklion Ocean SPAIN (Crete) Los Angeles Lisbon TURKEY UNITED Bermuda Ceuta Jerusalem/Bethlehem STATES (West End) (Spanish Morocco) Seville (Ashdod) ine (Cadiz) ISRAEL Athens e JORDAN Dubai Agadir (Piraeus) Aqaba Pacific MEXICO Madeira UNITED ARAB Ocean MOROCCO l Dat L (Funchal) Malta EMIRATES Ft. Lauderdale CANARY (Valletta) Suez Abu ISLANDS Canal Honolulu Huatulco Dhabi ne inn Puerto Santa Cruz Lanzarote OMAN a a Hawaii r o Hilo Vallarta NICARAGUA (Arrecife) de Tenerife Salãlah t t Kuala Lumpur I San Juan del Sur Cartagena (Port Kelang) Costa Rica COLOMBIA Sri Lanka PANAMA (Puntarenas) Equator (Colombo) Singapore Equator Panama Canal MALAYSIA INDONESIA Bali SAMOA (Benoa) AMERICAN Apia SAMOA Pago Pago AUSTRALIA South Pacific South Indian Ocean Atlantic Ocean Ocean Perth Auckland (Fremantle) Adelaide Sydney New Plymouth Burnie Picton Departure Ports Tasmania Christchurch More Ashore (Lyttelton) Overnight Fiordland NEW National Park ZEALAND up to continentscontinents (North America, South America, 111 51 Australia, Europe, Africa -

IUCN Evaluation of Nominations of Natural and Mixed Properties to the World Heritage List
WHC-02/CONF.201/INF.3 Convention Concerning the Protection of the World Cultural and Natural Heritage IUCN Evaluation of Nominations of Natural and Mixed Properties to the World Heritage List Report to the Bureau of the World Heritage Committee Twenty-sixth session 8-13 April 2002 - Paris, France Prepared by IUCN – The World Conservation Union 22 February 2002 Table of Contents 1. INTRODUCTION............................................................................................................................................iii 2. IUCN TECHNICAL EVALUATION REPORTS..........................................................................................1 A. Nominations of natural properties to the World Heritage List ...........................................................1 Pendjari and W National Parks (Benin) ...............................................................................................3 Rift Valley Lakes Reserve (Kenya)......................................................................................................5 Uvs Nuur Basin (Mongolia/Russian Federation)..................................................................................7 B. Nominations of mixed properties to the World Heritage List..............................................................9 Archipelago of La Maddalena (Italy) .................................................................................................11 i THE WORLD HERITAGE CONVENTION IUCN TECHNICAL EVALUATION REPORTS 22 February 2002 1. INTRODUCTION This technical