The British Lithostrotiontidae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Railway Ramblings
RAILWAY RAMBLINGS www.railwayramblers.org.uk SPRING 2011 No. 129 A TICK IN THE BOX INDICATES SUBSCRIPTION DUE OFFICERS Contact details have been removed to protect officers’ privacy in this sample edition published on the Internet Founder Nigel Willis. President David Shepherd, CBE Vice Presidents Bill Pertwee, MBE, and Paul Atterbury Chairman Richard Martin Vice Chairman Gordon Pirie Webmaster Jeff Vinter and Acting Editor Editor from Jonathan Dawson March 2011 Magazine Rupert Nicholson, Distribution Officer Treasurer Marion Thornton, Secretary Graham Lambert Membership Pete Walker Secretary AREA GROUP CONTACTS Chilterns Geoff Sargeant Eastern Phil Wood Midlands, East & West Temporary: Phillip Earnshaw Midlands, South Barry Bubb North East Ian Black North West Mark Jones Scotland Tony Jervis South West Vacant – volunteers please contact the Secretary Southern Graham Lambert – see under Secretary, above Wales Chris Parker Yorkshire Jane Ellis Enquiries Please address membership enquiries to the Membership Secretary and general enquiries to the Secretary. Contacting Officers No telephone calls after 9:30 pm, please. Kindly use email wherever possible Editorial I would like to begin by thanking David Brace for his efforts in producing the last five magazines, and for conducting a survey to find out what members actually wanted from this publication. This was worth doing, not least since such a survey was years overdue, and I have made a start at implementing his findings. While the magazine was being reviewed, all manner of suggestions were made, but I recommend that we concentrate first on getting the content right before looking at other possibilities such as a glossy finish and colour pictures. -

Palaeontological Society of Japan
Transactions and Proceedings of the Palaeontological Society of Japan New Series No. 81 Palaeontological Society of Japan April 20, 1971 Editor: Takashi HAMADA Associate edito r: Yasuhide IWASAKI Officers for 1971 - 1972 Honorary President : Teiichi KOB AYASHI President: Tokio SHIKAMA Councillors (* Executives): Kiyoshi ASANO*, Kiyotaka CHI NZEI*, Takashi HAMADA*, Tetsuro HA NA I*, Kotora HATAI, Itaru HAYAMI, Koichiro ICHIKAWA, Taro KANAYA, Kametoshi KA NMERA, Tamio KOTAKA, Tatsuro MATSUMOTO*, Hiroshi OZAKI*, Tokio SHIKAMA*, Fuyuji TAKAI *, Yokichi TAKAYANAGI Secretaries: Wataru HASHIMOTO, Saburo KA NNO Executive Committee General Affairs: Tetsuro HANAI, Naoaki AOKI Membership: Kiyotaka CHI NZEI, Toshio KOIKE Finance : Fuyuji T AKAI, Hisayoshi !Go Planning : Hiroshi OzAKI, Kazuo ASAMA Publications Transactions: Takashi HAMADA, Yasuhide IWASA KI Special Papers : Tatsuro MATSUMOTO, Tomowo 0ZA WA " Fossils": Kiyoshi ASANO, Toshiaki T AKAYAMA Fossil on the cover is left lower M2 of Pala eoloxodon naumanni (MAKIYAMA, 1924) from the uppermost part of the Tokyo formation (Upper Pleistocene) at Ikebukuro, Tokyo. All communications relating to this journal should be addressed to the PALAEONTOLOGICAL SOCIETY OF JAPAN c/o Geological Institute, Faculty of Science, University of Tokyo, Tokyo 113, Japan Sole agent: University of Tokyo Press, Hongo, Tokyo Trans. Proc. Palaeont. Soc. Japan, N.S., No. 81, pp. 1-10, pl. 1, April 20, 1971 574. J. FLEMING'S SPECIES OF BRITISH LOWER CARBONIFEROUS CORALS MAKOTO KATO Department of Geology and Mineralogy, Hokkaido University 7 v ~ ~ :7'E:Ncil&cv~OOJ11TJVJ:fii!J'Hcl!lllf/1lft:fi: 7 v ~ ~ :7'fi J:l!.:(£:t5::. tJ:b:ll.:·n,, ~rfli45 il;fc <b c. -?v'"C ~OOcv:fii!JHcllffi!Wl~~cli& L-t-::lrHJJcv ?f}cg-cv-A c: <0 ~, ji:WI'fi.:c. -

Directory of Resources
SETTLE – CARLISLE RAILWAY DIRECTORY OF RESOURCES A listing of printed, audio-visual and other resources including museums, public exhibitions and heritage sites * * * Compiled by Nigel Mussett 2016 Petteril Bridge Junction CARLISLE SCOTBY River Eden CUMWHINTON COTEHILL Cotehill viaduct Dry Beck viaduct ARMATHWAITE Armathwaite viaduct Armathwaite tunnel Baron Wood tunnels 1 (south) & 2 (north) LAZONBY & KIRKOSWALD Lazonby tunnel Eden Lacy viaduct LITTLE SALKELD Little Salkeld viaduct + Cross Fell 2930 ft LANGWATHBY Waste Bank Culgaith tunnel CULGAITH Crowdundle viaduct NEWBIGGIN LONG MARTON Long Marton viaduct APPLEBY Ormside viaduct ORMSIDE Helm tunnel Griseburn viaduct Crosby Garrett viaduct CROSBY GARRETT Crosby Garrett tunnel Smardale viaduct KIRKBY STEPHEN Birkett tunnel Wild Boar Fell 2323 ft + Ais Gill viaduct Shotlock Hill tunnel Lunds viaduct Moorcock tunnel Dandry Mire viaduct Mossdale Head tunnel GARSDALE Appersett Gill viaduct Mossdale Gill viaduct HAWES Rise Hill tunnel DENT Arten Gill viaduct Blea Moor tunnel Dent Head viaduct Whernside 2415 ft + Ribblehead viaduct RIBBLEHEAD + Penyghent 2277 ft Ingleborough 2372 ft + HORTON IN RIBBLESDALE Little viaduct Ribble Bridge Sheriff Brow viaduct Taitlands tunnel Settle viaduct Marshfield viaduct SETTLE Settle Junction River Ribble © NJM 2016 Route map of the Settle—Carlisle Railway and the Hawes Branch GRADIENT PROFILE Gargrave to Carlisle After The Cumbrian Railways Association ’The Midland’s Settle & Carlisle Distance Diagrams’ 1992. CONTENTS Route map of the Settle-Carlisle Railway Gradient profile Introduction A. Primary Sources B. Books, pamphlets and leaflets C. Periodicals and articles D. Research Studies E. Maps F. Pictorial images: photographs, postcards, greetings cards, paintings and posters G. Audio-recordings: records, tapes and CDs H. Audio-visual recordings: films, videos and DVDs I. -

\Yestliorland
188 'W"£lTIIY, \YESTliORLAND. W AITBY is a township, and under the provisions of SMARDALE, formerly a small township in the paris·h the "Local Government ...\et, 1894 " (56 and 57 Vict. c. of Kirkby Stephen, was, in 1895, under the provisions 73), was in 1895 formed into a parish, together with of the "Local Government Act, 1894.'' united for civil Smardale township, for civil purposes only, and is in purposes to the township of Waitby, and these two the :Northern division of the county, East Ward petty townships now form the parish of Wait by; it is 1 sessional division and union, county court district of mile west from Waitbv,. and 2/;- miles south-west from Appleby. The township is rl miles west-by-south from Kirl•by Stephen, with a st.ation on the North Eastern Kirkby Stephen, and 1 mile east from Smardale .station railway; the village, lying in a deep vale between Crosby on the North Eastern railway, and now (1914) consists and Ash fells, contains six houses only. Smardale Hall, of but nine houses, although it is supposed to have been now occupied as a farmhouse, is a mansion of the late in early times a market town, and possessed a castle, Tudor or Jacobean period, and was erected by the traces of which are still evident on Castle Carrick Hill, Dalstons, who came into possession of the manor by the e .1d between that eminence and the village traces of a marriage of Sir John Dalston kt. with Frances Warcop, chap~l have bePn IY'Pt with. -
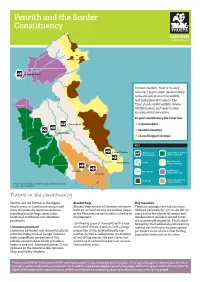
Penrith and the Border Constituency
Penrith and the Border Constituency Rockcliffe Marsh Cumbria Wildlife Trust is the only voluntary organisation devoted solely to the conservation of the wildlife and wild places of Cumbria. The Trust stands up for wildlife, creates wildlife havens, and seeks to raise environmental awareness. In your constituency the Trust has: Thacka Beck 2,219 members Eycott Hill 8 nature reserves Newton Reigny 2 Local Support Groups KEY Augill Pasture Wildlife Trust Lakeland Fells & Dales Nature Reserve Living Landscape North Pennines Coast Argill Woods Living Landscape Solway Basin Tarn Sike River/Stream Living Landscape Smardale Orton & Howgill Fells Peat bogs Living Landscape Crown Copyright and/or database right. All rights reserved. Licence no. 100044280 Nature in the constituency Penrith and the Borders is the biggest Blanket bogs Hay meadows constituency in Cumbria covering a vast Blanket bogs are one of the most extensive These increasingly rare habitats have array of nationally important habitats habitats in Cumbria and the shallow slopes declined nationally by 97% in the last 50 including blanket bogs, mires, lakes, of the Pennines are particularly suited to its years and so the cluster of species-rich heathland and flower-rich limestone development. meadows that still exist around Orton grasslands. are increasingly important. This habitat, Confined to areas of the world with a cool formed by the traditional practice of hay Limestone pavement and humid climate, Cumbria holds a large making, can hold up to 50 plant species Limestone pavement was formed by glacial proportion of this internationally rare per square metre and is a vital feeding scouring during the last ice age. -

Winter Weather on the S&C
Quarterly Magazine February 2016 No 143 JOURNAL Price £2.50 Winter weather on the S&C The Friends of the Settle - Carlisle Line FRIENDS OF THE SETTLE – CARLISLE LINE Settle Railway Station, Station Road, Settle, North Yorkshire BD24 9AA President: The Hon. Sir William McAlpine Bt. Vice Presidents: Lord Inglewood DL; The Bishop of Carlisle; Edward Album; Ron Cotton; Ann Cryer ; David Curry; Philip Johnston; Eric Martlew; Pete Shaw; Ken Shingleton; Brian Sutcliffe MBE; Gary Waller; David Ward. Chairman: Richard Morris - richard.morris @settle-carlisle.com Committee: Douglas Hodgins * (Vice-chairman & Futures Group Representative) [email protected] Stephen Way (Treasurer) [email protected] Paul Kampen (Secretary & Editor) [email protected] Peter Davies (Membership Secretary) [email protected] Ruth Evans (Volunteers Co-ordinator & Events Organiser) [email protected] John Johnson (Armathwaite signalbox & Carlisle representative) [email protected] Paul Levet (Business Development Co-ordinator) [email protected] Rod Metcalfe (On-train Guide Co-ordinator & Technology Adviser) [email protected] Phil Moorhouse (Insurance Consultant) [email protected] Pat Rand (Customer Relations, Trading & Settle Shop Manager) [email protected] Pete Shaw (Magazine Mailing Co-ordinator) Telephone 01274 590453 Nigel Ward (Solicitor) [email protected] * Indicates that these members were co-opted after the 2015 Annual General Meeting in accordance with the FoSCL constitution. Postal Addresses: Chairman: Richard Morris – 10 Mill Brow, Armathwaite, Carlisle CA4 9PJ Secretarial Enquiries, Hard Copy for the Magazine and General Postal Enquiries: Paul Kampen - 74 Springfield Road, Baildon, Shipley, W. -

Eden Valley Fringe
1 Valley of Dalebanks Beck, to the west of Crosby Ravensworth of Crosby the west Beck, to of Dalebanks Valley LCA 1 Eden Valley Fringe Yorkshire Dales National Park - Landscape Character Assessment YORKSHIRE DALES NATIONAL PARK LANDSCAPE CHARACTER ASSESSMENT LANDSCAPE CHARACTER AREAS 2 LCA 1 Eden Valley Fringe Key characteristics • Sheltered and enclosed farmland landscape on gently undulating slopes of the small tributary valleys of the River Eden which run north-east from the Orton Fells limestone plateau • The steep becks on the edge of the limestone plateau flow sporadically, but there is a dense network of small, winding streams within the valleys and throughout the lowland valley fringe landscape • Well settled landscape with spring-line villages on the edge of the fells to the south and nucleated stream-side villages within the valleys • Small-scale, historic field pattern on fringes of villages, with strong patterns of long tofts defined by walls, and isolated farmsteads • Fields are enclosed by limestone walls on the fringes of the limestone plateau and by a mixture of stone walls and hedgerows within the valleys. • Narrow tree lined gills and becks are perceptible in the more open upper valley landscapes as wooded features; small blocks of woodland, tree belts and hedgerow trees create a fairly enclosed landscape character overall • Small historic villages, built with local limestone and ‘pink’ sandstone, often centred on long greens • Villages are connected by a network of winding, narrow lanes, which are often enclosed by tall hedgerows. • Occasional long views from the upper valley slopes across the Eden Valley Fringe LCA to the upland ridges of the North Pennines and Cumbrian Fells to the north and west. -

Carboniferous Rugosa and Heterocorallia In
ACT A PAL A EON T 0 LOG I CAP 0 LON• ICA Vol. 22 1977 Nr. 4 NGUYEN DUC KHOA CARBONIFEROUS RUGOSA AND HETEROCORALLIA FROM BOREHOLES IN THE LUBLIN REGION (POLAND) Abstract. - Fourty two species referred to 21 genera and 2 subgenera are described; one genus and 5 species are new: Diphyphyllum rarevesiculosum sp.n., Lithostro tion (Siphonodendron) dobrolyubovae sp.n., L. (S.) rossicum strzelcense subsp.n., L. (S.) rossicum parvum subsp.n., Lublinophyllum fedorowskii gen. et sp.n. Genera Orionastraea and Turbinatocaninia are for the first time recorded in Poland. Hete rophyllidae with 5 septa and pentagonal cross-section are recorded. Blastogeny has been studied in 13 species and subspecies. Axial increase has been studied in detail in Lithostrotion (Siphonodendron) dobrolyubovae sp.n. Taxonomic division of Ru gosa based on blastogeny, is demonstrated to be questionable in some cases. Fusion of corallites has been observed in colonies of Lithostrotion and Lonsdaleia colonies. Existence of holotheca in Orionastraea aff. magna Kato & Mitchell has been proved. INTRODUCTION Carboniferous Rugosa and Heterocorallia in Polapd are known from the outcorps and boreholes in Sudetes, Silesia-Cracow Upland, Holy Cross Mts, Lublin region and Western Pomerania (Kunth 1869; Schindewolf 1941, 1942, 1952; Fedorowski 1968, 1970, 1971, 1975). Their stratigraphic and geographic distribution is shown in Table 1. In the Lublin region the Carboniferous system is represented by Visean, . Namurian, and Westfalian deposits. History of geological investigations in this region as well as descriptive palaeontology, lithology, and tectonics are presented in the studies by Bojkowski (1966a,b), Cebulak & Porzycki (1966), Porzycki (1976), and Zelichowski (1972c) among others. -

June 2011 Minutes
RAVENSTONEDALE PARISH COUNCIL Minutes of the Meeting held on Thursday 9 th June 2011 in Ravenstonedale High Chapel Room at 7.30pm Present Ernest Leach (Chairman) Cllrs Helen Shields, Hilary Law , Alison Morriss, James Richardson and Paul Capstick, Clerk. Chris Elphick One member of the public Apologies for absence had been received from County Councillor Peter Thornton, District Councillor Ian Torkington and UECP Project Officer Libby Bateman. Declarations of Interest Cllr Law declared an interest in the discussions on the Scandal Beck wall renovations. She is a partner in the construction company carrying out the work. Cllr Morriss declared an interest in Planning Application 11/0420 and 11/0423. She is a partner in the company carrying out the work. Cllr Morris also declared an interest in the proposed signage at Scandal Beck Green as her company may tender for the work. Cllr Morriss further declared an interest in Planning Application No. 11/0328. She is one of the applicants. Minutes of the Annual General Meeting held on Thursday 12 th May 2011 had been circulated and were signed by the Chairman as a correct record. Co-option of Parish Councillor Only six nominations were received at the recent Parish Council Elections for the seven seats on Ravenstonedale Parish Council. The vacancy may now be filled without the need for further election. At the AGM held on 12 th May it had been resolved that the vacancy be advertised in order to ensure equal opportunity for all. There was one applicant, Mr Alex Watts- Tobin from Newbiggin-on-Lune. Mr Watts-Tobin is known to all councillors. -

Late Carboniferous Colonial Rugosa (Anthozoa) from Alaska
Geologica Acta, Vol.12, Nº 3, September 2014, 239-267 DOI: 10.1344/GeologicaActa2014.12.3.6 Late Carboniferous colonial Rugosa (Anthozoa) from Alaska J. FEDOROWSKI1 and C.H. STEVENS2 1Institute of Geology, Adam Mickiewicz University Maków Polnych 16, PL-61-606, Poznań, Poland. E-mail: [email protected] 2 Department of Geology, San José University San José, California 95192, USA. E-mail: [email protected] ABS TRACT Late Carboniferous colonial corals from the Moscovian Saginaw Bay Formation and the underlying Bashkirian crinoidal limestone exposed on northeastern Kuiu Island and a nearby islet, part of the Alexander terrane in southeastern Alaska, are described and illustrated for the first time, and are supplemented by revision, redescription and reillustration of most Atokan specimens from Brooks Range, northern Alaska, first described by Armstrong (1972). New taxa from the Kuiu Island area include the new species Paraheritschioides katvalae and the new genus and species Arctistrotion variabilis, as well as the new Subfamily Arctistrotioninae. The corals Corwenia jagoensis and Lithostrotionella wahooensis of Armstrong (1972) also are redefined and redescribed. Paraheritschioides jagoensis is based on the holotype of ‘C’. jagoensis. P. compositus sp. nov. is based on a “paratype” of ‘C.’ jagoensis. In addition to a redefinition and redescription of ‘L.’ wahooensis as Arctistrotion wahooense, one “paratype” of that species is described as A. simplex sp. nov. The phylogeny and suspected relationships of some fasciculate Carboniferous Rugosa also are discussed. Based on relationships and similarities within the Late Carboniferous colonial Rugosa from the Brooks Range, Kuiu Island and the eastern Klamath terrane, we conclude that all three areas were geographically close enough at that time so that larvae were occasionally dispersed by oceanic currents. -

Appleby Kirkby St
Grades Bikes on trains Cycle shops and hire Rides are graded by distance Northern Rail runs the trains on the The nearest bike shop is Kirkby Stephen Cycle Centre (short, medium, long, extra long) Leeds-Settle-Carlisle line. Although in Kirkby Stephen (Station Yard) who hire out mountain and height gain per kilometre strictly speaking it’s only two bikes per bikes - 017683 71658 (easy, moderate, hard, challenge, train, the conductors will do their best extreme). Short cuts are shown on to get you and your bike on board. Stonetrail Holidays near Ravenstonedale (Street Farm) some rides. Timetable and other information is on has mountain bikes for hire - 015396 23444 the website www.settle-carlisle.co.uk Good Cycling Code Following the routes Tourist Information Centres (TIC) Each route is highlighted on its own Enjoy a 15-mile (24km) linear ride Obey the rules of the road Appleby TIC Ordnance Survey map with arrows from Kirkby Stephen station to • Always follow the Highway Code Explore the Upper Eden Valley’s Moot Hall, Boroughgate, Appleby CA16 6XE showing which way round to go and Appleby station following the blue • Ride in single file to let vehicles overtake added directions at numbered points 71 and 20 signs (from the town Mountain biking 017683 51177 delightful mix of scenery and Be courteous [email protected] to help with navigation. The ‘Key to centre) to Soulby and Whygill Head, There are numerous byway and • Don’t assume everyone can see or hear you maps / route descriptions’ explains the then 71 and 68 signs through Great Kirkby Stephen TIC bridleways tracks around Kirkby • Give way to pedestrians, wheelchair users heritage at a slower pace symbols shown on the maps and the Asby to Appleby. -

Chapter 5. Paleozoic Invertebrate Paleontology of Grand Canyon National Park
Chapter 5. Paleozoic Invertebrate Paleontology of Grand Canyon National Park By Linda Sue Lassiter1, Justin S. Tweet2, Frederick A. Sundberg3, John R. Foster4, and P. J. Bergman5 1Northern Arizona University Department of Biological Sciences Flagstaff, Arizona 2National Park Service 9149 79th Street S. Cottage Grove, Minnesota 55016 3Museum of Northern Arizona Research Associate Flagstaff, Arizona 4Utah Field House of Natural History State Park Museum Vernal, Utah 5Northern Arizona University Flagstaff, Arizona Introduction As impressive as the Grand Canyon is to any observer from the rim, the river, or even from space, these cliffs and slopes are much more than an array of colors above the serpentine majesty of the Colorado River. The erosive forces of the Colorado River and feeder streams took millions of years to carve more than 290 million years of Paleozoic Era rocks. These exposures of Paleozoic Era sediments constitute 85% of the almost 5,000 km2 (1,903 mi2) of the Grand Canyon National Park (GRCA) and reveal important chronologic information on marine paleoecologies of the past. This expanse of both spatial and temporal coverage is unrivaled anywhere else on our planet. While many visitors stand on the rim and peer down into the abyss of the carved canyon depths, few realize that they are also staring at the history of life from almost 520 million years ago (Ma) where the Paleozoic rocks cover the great unconformity (Karlstrom et al. 2018) to 270 Ma at the top (Sorauf and Billingsley 1991). The Paleozoic rocks visible from the South Rim Visitors Center, are mostly from marine and some fluvial sediment deposits (Figure 5-1).