Hybridization Between Asclepias Purpurascens and Asclepias Syriaca (Apocynaceae): a Cause for Concern?1
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bwsr Featured Plant Minnesota's Milkweeds
BWSR FEATURED PLANT MINNESOTA’S MILKWEEDS Publication Date: 6‐1‐13 Milkweeds play a key role in wetlands, prairies, savannas and forests in Minnesota. The genus (Asclepias) is particularly important as a nectar and larval food source for a wide range of insect species. The best known example is the monarch butterfly whose larvae appear to feed only on milkweeds. Milkweeds have a unique pollination mechanism where pollen grains are enclosed in waxy sacs called “pollina” that attach to the legs of butterflies, moths, bees, ants and wasps and are then deposited in another milkweed flower if they step into a specialized anther opening. Most milkweeds are toxic to vertebrate herbivores due to cardiac glycosides that are in their plant cells. In addition to supporting insect populations, Butterfly Milkweed milkweeds also provide other landscape benefits due to their extensive root systems (sometimes deep roots, sometimes horizontal) that Photos by Dave Hanson decrease compaction, add organic material to the soil and improve unless otherwise stated water infiltration. Common milkweed is probably the best known milkweed species as it is found in all counties of the state and was included on some county prohibited noxious weed lists. The species was considered a common agricultural weed as its extensive root network made it difficult to remove from agricultural fields with cultivators. Now the species is effectively removed from genetically modified corn and soybean fields that are sprayed with herbicide. This practice has contributed to significant declines in milkweed species, with an estimated 58% decline in the Midwest between 1999 and 2010 and a corresponding 81% decline in monarch butterfly production (Pleasants & Oberhauser, 2013). -

Asclepias Purpurascens L. Purple Milkweed
Asclepias purpurascens L. purple milkweed State Distribution Photo by Michael R. Penskar Best Survey Period Jan Feb Mar Apr May Jun Jul Aug Sept Oct Nov Dec Legal status: State threatened one or two additional umbels are present in the upper leaf axils. The individual flowers, which are usually Global and state rank: G4G5/S3 from 13-17 mm long, bear reflexed, purplish corolla lobes that are glabrous (smooth), pale purple hoods Family: Asclepiadaceae (milkweed family) (forming the corona) 5-7 mm long, and incurved flat horns that are shorter than the hood. The reproductive Total range: Asclepias purpurascens is found parts (filaments, anthers, and style) are fused into a principally in eastern North America, occurring from structure called the gynostegium. The fruit is a smooth New Hampshire south to Virginia and ranging west to follicle (a pod) filled with seeds attached to downy hairs Wisconsin, Iowa, Kansas, and Oklahoma. (coma) that aid in wind dispersal. State distribution: Purple milkweed is known from Asclepias purpurascens is often difficult to distinguish more than 60 occurrences in southern Michigan; thirty- from the very similar looking common milkweed, four of these records are derived from collections made Asclepias syriaca, which despite its unfortunate prior to 1930. This species is concentrated primarily Latin epithet is also a native milkweed. Overall, the in southeastern and southwestern Lower Michigan, leaves of A. purpurascens are more acute and less where it is known from 19 counties, with most counties predominately pinnately–veined (i.e. more strongly tallying only a single occurrence. Counties with the net-veined) than A. -

Natural Communities of Michigan: Classification and Description
Natural Communities of Michigan: Classification and Description Prepared by: Michael A. Kost, Dennis A. Albert, Joshua G. Cohen, Bradford S. Slaughter, Rebecca K. Schillo, Christopher R. Weber, and Kim A. Chapman Michigan Natural Features Inventory P.O. Box 13036 Lansing, MI 48901-3036 For: Michigan Department of Natural Resources Wildlife Division and Forest, Mineral and Fire Management Division September 30, 2007 Report Number 2007-21 Version 1.2 Last Updated: July 9, 2010 Suggested Citation: Kost, M.A., D.A. Albert, J.G. Cohen, B.S. Slaughter, R.K. Schillo, C.R. Weber, and K.A. Chapman. 2007. Natural Communities of Michigan: Classification and Description. Michigan Natural Features Inventory, Report Number 2007-21, Lansing, MI. 314 pp. Copyright 2007 Michigan State University Board of Trustees. Michigan State University Extension programs and materials are open to all without regard to race, color, national origin, gender, religion, age, disability, political beliefs, sexual orientation, marital status or family status. Cover photos: Top left, Dry Sand Prairie at Indian Lake, Newaygo County (M. Kost); top right, Limestone Bedrock Lakeshore, Summer Island, Delta County (J. Cohen); lower left, Muskeg, Luce County (J. Cohen); and lower right, Mesic Northern Forest as a matrix natural community, Porcupine Mountains Wilderness State Park, Ontonagon County (M. Kost). Acknowledgements We thank the Michigan Department of Natural Resources Wildlife Division and Forest, Mineral, and Fire Management Division for funding this effort to classify and describe the natural communities of Michigan. This work relied heavily on data collected by many present and former Michigan Natural Features Inventory (MNFI) field scientists and collaborators, including members of the Michigan Natural Areas Council. -

Identification of Milkweeds (Asclepias, Family Apocynaceae) in Texas
Identification of Milkweeds (Asclepias, Family Apocynaceae) in Texas Texas milkweed (Asclepias texana), courtesy Bill Carr Compiled by Jason Singhurst and Ben Hutchins [email protected] [email protected] Texas Parks and Wildlife Department Austin, Texas and Walter C. Holmes [email protected] Department of Biology Baylor University Waco, Texas Identification of Milkweeds (Asclepias, Family Apocynaceae) in Texas Created in partnership with the Lady Bird Johnson Wildflower Center Design and layout by Elishea Smith Compiled by Jason Singhurst and Ben Hutchins [email protected] [email protected] Texas Parks and Wildlife Department Austin, Texas and Walter C. Holmes [email protected] Department of Biology Baylor University Waco, Texas Introduction This document has been produced to serve as a quick guide to the identification of milkweeds (Asclepias spp.) in Texas. For the species listed in Table 1 below, basic information such as range (in this case county distribution), habitat, and key identification characteristics accompany a photograph of each species. This information comes from a variety of sources that includes the Manual of the Vascular Flora of Texas, Biota of North America Project, knowledge of the authors, and various other publications (cited in the text). All photographs are used with permission and are fully credited to the copyright holder and/or originator. Other items, but in particular scientific publications, traditionally do not require permissions, but only citations to the author(s) if used for scientific and/or nonprofit purposes. Names, both common and scientific, follow those in USDA NRCS (2015). When identifying milkweeds in the field, attention should be focused on the distinguishing characteristics listed for each species. -

Physiological and Chemical Studies Upon the Milkweed (Asclepias Syriaca L) Fisk Gerhardt Iowa State College
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 1928 Physiological and chemical studies upon the milkweed (Asclepias syriaca L) Fisk Gerhardt Iowa State College Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Agricultural Science Commons, Agriculture Commons, and the Plant Biology Commons Recommended Citation Gerhardt, Fisk, "Physiological and chemical studies upon the milkweed (Asclepias syriaca L)" (1928). Retrospective Theses and Dissertations. 14748. https://lib.dr.iastate.edu/rtd/14748 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMl films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMl a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overiaps. -

Risk Assessment of Asclepias Syriaca
Risk Assessment of Asclepias syriaca Name of Organism: Asclepias syriaca L. – common milkweed Objective: Assess the risks associated with this species in EU Version: NAPRA EU amendment Final 30/11/2015 Author(s) Barbara Tokarska-Guzik1, Ewa Pisarczyk2 Expert reviewer Zoltán Botta-Dukát3, Jan Pergl4 Notes: Confidence is rated as low, medium, high or very high. Likelihood is rated as very unlikely, unlikely, moderately likely, likely or very likely. The percentage categories are 0% - 10%, 11% - 33%, 34% - 67%, 68% - 90% or 91% - 100%. N/A = not applicable. 1 University of Silesia in Katowice 2 General Directorate for Environmental Protection in Poland 3 Center for Ecological Research, Hungarian Academy of Sciences 4 Institute of Botany, Academy of Sciences of the Czech Republic Page 1 of 26 EU CHAPPEAU QUESTION RESPONSE 1. In how many EU member states has this species been recorded? List them. 17: Austria, Belgium, Bulgaria, Croatia, Czech Republic, Denmark, France, Germany, Hungary, Italy, Lithuania, Netherlands, Poland, Romania, Slovakia, Slovenia, Spain, Sweden Sanz-Elorza et al. 2001; Essl and Rabitsch 2002, 2004; Tokarska-Guzik 2005; Verloove 2006; Bagi 2008; Boršić et al. 2008; Jogan et al. 2012; Medvecká et al. 2012; Pyšek et al. 2012; Petrova et al. 2013; Mitić 2013; DAISIE 2015; FCD 2015; NOBANIS 2015; Q-bank 2015; http://svenskbotanik.se; Zimmermann et al. 2015 2. In how many EU member states has this species currently established 13: Austria, Bulgaria, Croatia, Czech Republic, Denmark, France, Hungary, Italy, Lithuania, populations? List them. Netherlands, Poland, Romania, Slovakia CABI 2011, DAISIE 2015; NOBANIS 2015; 3. In how many EU member states has this species shown signs of 11: Austria, Bulgaria, Croatia, Czech Republic, Hungary, Italy, Lithuania, Netherlands, Poland, invasiveness? List them. -

Open Myers THESIS.Pdf
The Pennsylvania State University The Graduate School School of Science, Engineering, and Technology IMPACT OF OZONE ON MILKWEED (ASCLEPIAS) SPECIES A Thesis in Environmental Pollution Control by Abigail C. Myers 2016 Abigail C. Myers Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science December 2016 The thesis of Abigail C. Myers was reviewed and approved* by the following: Dennis R. Decoteau Professor of Horticulture and Plant Ecosystem Health Thesis Advisor Donald D. Davis Professor of Plant Pathology and Environmental Microbiology Richard Marini Professor of Horticulture Shirley Clark Associate Professor of Environmental Engineering Graduate Program Coordinator, Environmental Engineering and Environmental Pollution Control *Signatures are on file in the Graduate School ii Abstract Tropospheric (or ground level) ozone in ambient concentrations can damage vegetation and interfere with the human respiratory system. Plants as bioindicators of ozone are commonly used to detect phytotoxic levels of tropospheric ozone where physical/chemical/electrical monitoring equipment cannot be utilized due to expense, electrical needs, or availability of instruments. Asclepias syriaca (Common Milkweed) has been effectively used as a bioindicator for ozone. Visual ozone injury on Common Milkweed is characterized as purple stippling of the upper surface of older leaves as the season progresses, the purple coloration of the upper leaf surface may encompass most of the leaf surface. While sensitivity to ozone has been documented on Common Milkweed, less is known about the ozone sensitivity of other Asclepias species and little is known regarding the concentration dose response of Common Milkweed to ozone and timing of visual symptoms. Of the Asclepias species evaluated Tropical Milkweed (A. -
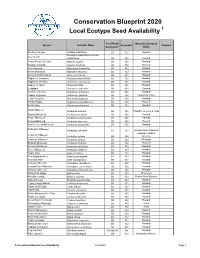
Local Ecotype Seed Species Available
Conservation Blueprint 2020 Local Ecotype Seed Availability 1 Seed Origin Nebraska County of Species Scientific Name Ecoregion Request Generation 2 Origin Western Yarrow Achillea millefolium G0 332 Howard Acmispon americanus or Lotus Deer Vetch unifoliolatus G0 332 Howard Rough Purple Gerardia Agalinis aspera G0 332 Howard Slender Gerardia Agalinis tenuifolia G0 332 Howard Anise Hyssop Agastache foeniculum G2 332 Howard White Snakeroot Ageratina altissima G0 332 Howard Wild Onion/Wild Garlic Allium canadense G0 332 Howard Ragweed, Common Ambrosia artemisiifolia G0 332 Howard Ragweed, Western Ambrosia psilostachya G0 332 Howard Ragweed, Giant Ambrosia trifida G0 332 Howard Leadplant Amorpha canescens G0 332 Howard Caroline Anemone Anemone caroliniana G0 332 Howard Candle Anemone Anemone cylindrica G0 332 Howard & Colfax Field Pussytoes Antennaria neglecta G0 332 Howard Prickly Poppy Argemone polyanthemos G0 332 Howard White Sage Artemisia ludoviciana G0 332 Howard Sand Milkweed Asclepias arenaria G0 332 Howard, Greeley & Loup Swamp Milkweed Asclepias incarnata G1 332 Howard Purple Milkweed Asclepias purpurascens G1 251 Kansas Showy Milkweed Asclepias speciosa G1 332 Howard Narrow-leaved Milkweed Asclepias stenophylla G0 332 Howard Sullivant's Milkweed Asclepias sullivantii G1 332 Howard, Knox & Merrick Howard, Colfax & Common Milkweed Asclepias syriaca G0 332 Greeley Common Milkweed Asclepias syriaca G0 222 Michigan Butterfly Milkweed Asclepias tuberosa G1 332 Howard Whorled Milkweed Asclepias verticillata G0 332 Howard Green Milkweed -

MONARCHS in PERIL HE MONARCH BUTTERFLY IS in SERIOUS TROUBLE —Their Numbers Have Tplummeted Over the Past Two Decades
MONARC HS IN PERIL HERBICIDE-RESISTANT CROPS AND THE DECLINE OF MONARCH BUTTERFLIES IN NORTH AMERICA EXECUTIVE SUMMARY FEBRUARY 2015 ABOUT CENTER FOR FOOD SAFETY CENTER FOR FOOD SAFETY (CFS) is a non-profit public interest and environmental advocacy membership organization established in 1997 for the purpose of challenging harmful food production technologies and promoting sustainable alternatives. CFS combines multiple tools and strategies in pur suing its goals, including litigation and legal petitions for rulemaking, legal support for various sustainable agriculture and food safety constituencies, as well as public education, grassroots organizing and media outreach. ACKNOWLEDGEMENTS Authors: BILL FREESE AND MARTHA CROUCH, P hD Executive Summary Contributing Writer: LARISSA WALKER Copy Editing: ABIGAIL SEILER, LARISSA WALKER, MADELEINE CARNEMARK Legal Consultant: GEORGE KIMBRELL Graphics: PATRICK RIGGS Design: HUMMINGBIRD DESIGN STUDIO Report Advisor: ANDREW KIMBRELL The authors are indebted to several reviewers, in particular Dr. Lincoln Brower, for their helpful comments and suggestions. CENTER FOR FOOD SAFETY MONARCHS IN PERIL HE MONARCH BUTTERFLY IS IN SERIOUS TROUBLE —their numbers have Tplummeted over the past two decades. The butterfly’s decline tracks the virtual eradication of its caterpillar’s chief food source—common milkweed—from Midwestern cropland. The demise of milkweed is due to intensive spraying of glyphosate herbicide on Monsanto’s Roundup Ready corn and soybeans that have been genetically engi - neered to withstand it. Monarchs are in imminent danger unless milkweed is restored to Midwestern crop fields. Milkweed cannot recover with continued heavy use of glyphosate on Roundup Ready crops. We face a historic choice: do we want to protect Monsanto or mon - archs? The threats to monarch survival will soon escalate, if new genetically engineered (GE) crops resistant to glyphosate and additional herbicides like 2,4-D and dicamba are introduced. -

Milkweed and Monarchs
OHIO DEPARTMENT OF NATURAL RESOURCES DIVISION OF WILDLIFE MILKWEEDS AND MONARCHS Acknowledgments Table of Contents We thank Dr. David Horn, past president of the Ohio Lepidopterists, 03 MONARCH LIFE CYCLE for his thoughtful review of this publication. Our appreciation goes 04 MONARCH MIGRATION to the Ohio Lepidopterists, and Monarch Watch. These organizations work tirelessly to promote the conservation of butterflies and moths. 05 PROBLEMS & DECLINE COVER PHOTO BY KELLY NELSON 06 MILKWEEDS 07 OTHER MILKWEED SPECIALISTS 08 MONARCH NURSERY GARDEN Introduction 09 FIVE EXCELLENT MILKWEEDS Text and photos by Jim McCormac, Ohio Division of Wildlife, unless otherwise stated. 10 SUPERB MONARCH NECTAR SOURCES The Monarch butterfly (Danaus plexippus) is one of North Amer- ica’s most iconic insects. The gorgeous golden-brown and black but- terfly is probably the most celebrated insect on the continent, and the migration of the eastern population is conspicuous and spectacular. Southbound Monarchs can appear anywhere, even in highly urban- ized locales, and the butterflies often use backyard gardens as way sta- tions. Occasionally a resting swarm of hundreds or even thousands of butterflies is encountered. The spectacle of trees dripping with living leaves of butterflies is unlikely to be forgotten. PHOTO BY CHRIS FROST A Pictorial Journey From Caterpillar to Chrysalis to Butterfly PHOTOS BY STEVEN RUSSEL SMITH Monarch Butterfly Life Cycle Like all species in the order Lepidoptera (moths and butterflies), soon hatch. The caterpillars begin eating the milkweed foliage, and Monarchs engage in complete metamorphosis. This term indicates grow rapidly. The growth process involves five molts where the cater- that there are four parts to the life cycle: egg, caterpillar, pupa, and pillar sheds its skin and emerges as a larger animal. -

(Asclepias Syriaca) Occurrence in Iowa Cropland from 1999 to 2009 Robert G
Agronomy Publications Agronomy 12-2010 Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009 Robert G. Hartzler Iowa State University, [email protected] Follow this and additional works at: http://lib.dr.iastate.edu/agron_pubs Part of the Agricultural Science Commons, Agronomy and Crop Sciences Commons, Entomology Commons, and the Weed Science Commons The ompc lete bibliographic information for this item can be found at http://lib.dr.iastate.edu/ agron_pubs/33. For information on how to cite this item, please visit http://lib.dr.iastate.edu/ howtocite.html. This Article is brought to you for free and open access by the Agronomy at Iowa State University Digital Repository. It has been accepted for inclusion in Agronomy Publications by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. Reduction in common milkweed (Asclepias syriaca) occurrence in Iowa cropland from 1999 to 2009 Abstract The or le of common milkweed in the lifecycle of the monarch butterfly ash increased interest in the presence of this weed in the north central United States. An initial survey conducted in 1999 found that low densities of common milkweed occurred in approximately 50% of Iowa corn and soybean fields. In 2009, common milkweed was present in only 8% of surveyed fields, and the area within infested fields occupied by common milkweed was reduced by approximately 90% compared to 1999. The widespread adoption of glyphosate resistant corn and soybean cultivars and the reliance on post-emergence applications of glyphosate for weed control in crop fields likely has contributed to the decline in common milkweed in agricultural fields. -

Native Plants & Pollinators
Native Plants & Pollinators What is a “Native” plant? A plant that is a part of the balance of nature that has developed over hundreds or thousands of years in a particular region or ecosystem. What is Ecotype? “Ecotype” refers to a genetically unique population of plants adapted to a local environment. This matters when it comes to foliage, flowering, seed set, seed germination and overall plant performance. What is Nectar? Nectar is a sweet liquid secreted by plants to attract pollinators. It is an important food source for butterflies, bees and hummingbirds. What is Pollen? Pollen carries cells which enable fertilization of plants, thus enabling the plant to reproduce. Examples of Pollinators: Bees, butterflies, beetles, flies, as well as some birds and bats are pollinators. About 12% of the world’s flowering plants are wind-pollinated, including grasses, cereal crops, and many trees. Cross-pollination Self-pollination Why is Pollination So Important? Pollination is mutually beneficial to plants AND to pollinators. Pollination results in the production of seeds and is necessary for many plants to reproduce. Why is Pollination So Important? (cont’d) Pollinators receive nectar and/or pollen rewards from the flowers they visit. Sugary nectar provides pollinators with carbohydrates, while pollen offers proteins, fats, vitamins, minerals, and necessary phytochemicals. Why Do Humans Need Pollinators? Pollinators are necessary for three- quarters of our major food crops. Without pollinators, our diets would be severely limited, and it would be more difficult to acquire the variety of vitamins and minerals we need to stay healthy. Go Native! Add Native Plants Pollinators have evolved with native plants, which often need less water than ornamentals.