The Initial Phase of a Longleaf Pine-Wiregrass Savanna Restoration: Species Establishment and Community Responses
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Awards Ceremony
South’s BEST 2019 Final Results BEST Award Winners: 1st Place: 966 Starkville High School (Mississippi BEST) 2nd Place: 1351 Ridgecrest Christian Academy (Wiregrass BEST) 3rd Place: 607 Eastwood/Cornerstone Schools (Montgomery BEST) Game Winners: 1st Place Robotics: 966 Starkville High School (Mississippi BEST) 2nd Place Robotics: 873 MACH (Jubilee BEST) 3rd Place Robotics: 803 DARC (Tennessee Valley BEST) 4th Place Robotics (finalist): 1351 Ridgecrest Christian Academy (Wiregrass BEST) Middle School Awards: Top ranking middle school in the BEST Award competition: Woodlawn Beach Middle School (Team 1112 - Emerald Coast BEST) Top ranking middle school in the Robotics competition: Semmes Middle School (Team 881 – Jubilee BEST) The Briggs and Stratton Teacher Leadership Award • Kevin Welch from 1042 Stewarts Creek Middle School (Music City BEST) • Amy Sterling from 1564 Moulton Middle/Lawrence Co. High School (Northwest Alabama BEST) • Beth Lee from 1805 Davis-Emerson Middle School (Shelton State BEST) South’s BEST Volunteer Award • Abigail Madden • Kate Kramer • Beth Rominger (Shelton State BEST) Jim Westmoreland Memorial Judge’s Award • Louis Feirman (Jubilee BEST) 1 BEST Award Category Awards: Best Spirit and Sportsmanship Award: 1st Team: 867 Faith Academy (Jubilee BEST) 2nd Team: 1552 Brooks High School (Northwest Alabama BEST) 3rd Team: 966 Starkville High School (Mississippi BEST) Best Engineering Notebook Award: 1st Team: 851 W.P. Davidson High School (Jubilee BEST) 2nd Team: 1653 Fort Payne High School (Northeast Alabama BEST) 3rd -

Zone 3 – Atlanta Regional Commission
REGIONAL PROFILE ZONE 3 – ATLANTA REGIONAL COMMISSION TABLE OF CONTENTS ZONE POPULATION ........................................................................................................ 2 RACIAL/ETHNIC COMPOSITION ..................................................................................... 2 MEDIAN ANNUAL INCOME ............................................................................................. 3 EDUCATIONAL ACHIEVEMENT ...................................................................................... 4 GEORGIA COMPETITIVENESS INITIATIVE REPORT .................................................... 10 RESOURCES .................................................................................................................. 11 This document is available electronically at: http://www.usg.edu/educational_access/complete_college_georgia/summit ZONE POPULATION 2011 Population 4,069,211 2025 Projected Population 5,807,337 Sources: U.S. Census, American Community Survey 2011 ACS Demographic and Housing Estimates, 5-year estimate Georgia Department of Labor, Area Labor Profile Report 2012 RACIAL/ETHNIC COMPOSITION Source: U.S. Census, American Community Survey 2011 ACS Demographic and Housing Estimates, 5-year estimate 2 MEDIAN ANNUAL INCOME Source: U.S. Census, American Community Survey 2010, Selected Economic Characteristics, 5-year estimate 3 EDUCATIONAL ACHIEVEMENT HIGH SCHOOL GRADUATION RATES SYSTEM NAME 2011 GRADUATION RATE (%) Decatur City 88.40 Buford City 82.32 Fayette 78.23 Cherokee 74.82 Cobb 73.35 Henry -

Of the Wiregrass Primitive Baptists of Georgia: a History of the Crawford Faction of the Alabaha River Primitive Baptist Association, 18422007
The “Gold Standard” of the Wiregrass Primitive Baptists of Georgia: A History of the Crawford Faction of the Alabaha River Primitive Baptist Association, 18422007 A Thesis submitted to the Graduate School Valdosta State University in partial fulfillment of requirements for the degree of MASTER OF ARTS in History in the Department of History of the College of the Arts July 2008 Michael Otis Holt BAS, Valdosta State University, 2003 © 2008 Michael Otis Holt All Rights Reserved This thesis, “The ‘Gold Standard’ of the Wiregrass Primitive Baptists of Georgia: A History of the Crawford Faction of the Alabaha River Primitive Baptist Association, 18422007,” by Michael Otis Holt is approved by: Major Professor ___________________________________ John G. Crowley, Ph.D. Associate Professor of History Committee Members ____________________________________ Melanie S. Byrd, Ph.D. Professor of History ____________________________________ John P. Dunn, Ph.D. Assistant Professor of History _____________________________________ Michael J. Stoltzfus, Ph.D. Professor of Philosophy and Religious Studies Dean of Graduate School _____________________________________ Brian U. Adler, Ph.D. Professor of English Fair Use This thesis is protected by the Copyright Laws of the United States (Public Law 94553, revised in 1976). Consistent with fair use as defined in the Copyright Laws, brief quotations from this material are allowed with proper acknowledgement. Use of the material for financial gain without the author’s expressed written permission is not allowed. Duplication I authorize the Head of Interlibrary Loan or the Head of Archives at the Odum Library at Valdosta State University to arrange for duplication of this thesis for educational or scholarly purposes when so requested by a library user. -
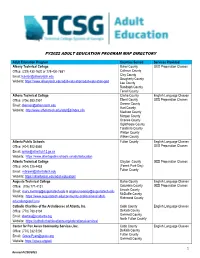
Fy2022 Adult Education Program Map Directory
FY2022 ADULT EDUCATION PROGRAM MAP DIRECTORY Adult Education Program Counties Served Services Provided Albany Technical College Baker County GED Preparation Classes Office: (229) 430-1620 or 229-430-7881 Calhoun County Email: [email protected] Clay County Dougherty County Website: https://www.albanytech.edu/adult-education/adult-education-ged Lee County Randolph County Terrell County Athens Technical College Clarke County English Language Classes Office: (706) 583-2551 Elbert County GED Preparation Classes Email: [email protected] Greene County Hart County Website: http://www.athenstech.edu/adultEd/index.cfm Madison County Morgan County Oconee County Oglethorpe County Taliaferro County Walton County Wilkes County Atlanta Public Schools Fulton County English Language Classes Office: (404) 802-3560 GED Preparation Classes Email: [email protected] Website: https://www.atlantapublicschools.us/adulteducation Atlanta Technical College Clayton County GED Preparation Classes Office: (404) 225-4433 (Forest Park-Day) Email: [email protected] Fulton County Website: https://atlantatech.edu/adult-education/ Augusta Technical College Burke County English Language Classes Office: (706) 771-4131 Columbia County GED Preparation Classes Email: [email protected] & [email protected] Lincoln County McDuffie County Website: https://www.augustatech.edu/community-and-business/adult- Richmond County educationgedell.cms Catholic Charities of the Archdiocese of Atlanta, Inc. Cobb County English Language Classes Office: -

John Crews of Camden and His Ancestors
John Crews of Camden and His Ancestors John Crews was born about 1763 in Virginia (1). He is a son of Stanley and Agnes Crews and a grandson to David and Mary Stanley Crews of Virginia (2). He was married to 1st Elizabeth aka Betsey (maiden name unknown); 2nd to Elizabeth Stafford Johns, widow of Jacob Johns. He was one of two of the surname Crews to settle in Camden County, Georgia, during the early-to-mid 1790s. The other Crews family being Isaac Crews, later the Clerk of Court in Camden County, Georgia (no relationship established). These two gentlemen were the first of the Crews surname to settle in Southeast Georgia. John’s grandfather, David Crews (born about 1710-1766) married Mary Stanley (1706- 1766) at the “Friends Meeting House” in Hanover County, Virginia on 9 Nov 1733/34 (2). The children attributed to this union of David and Mary (2) are: David Milton 2 Mar 1740 New Kent County, VA d. 2 Nov,1821 Madison Co., KY Elizabeth born about 1745 Hanover County, VA; m 3 July 1753 (3). John Crews Mary Crews m. Charles Ballew, died about 1811 Stanley - 1740 in VA, died 1792 Wilkes Co., GA m. Agnes (Martin) John's father, Stanley, was the only one of the above-mentioned siblings to settle into Georgia. All others appear to settle in Kentucky. The Stanley Crews family removed from Virginia to Wilkes County, Georgia, after the children were born and he and his brother David’s service in the Continental Army during the American Revolution (2). -

Georgia Board of Examiners of Licensed Practical Nurses
Georgia Board of Examiners of Licensed Practical Nurses, Approved Practical Nursing Programs Technical Program Contact Information Other Campus Approval NCLEX-PN 5- Year 2010 Average PN Programs Status College Director NCLEX Parent Institution Score 2006 - 2010 Albany Mattie Buchannon, 1704 South Slappey Boulevard FULL Candidates Candidates RN, MSN Technical Albany, Georgia 31701 34 / 34 Pass 155 /146 [email protected] College 100 % www.albanytech.edu 94.19 % (229) 430-1721 Altamaha Sheryl Nipper, RN, 1777 West Cherry Street Baxley Campus FULL Candidates Candidates BSN Technical Jesup, Georgia 31545 41 / 39 Pass 164 / 149 [email protected] College Golden Isles Career 95.12 % www.altamahatech.edu Pass 90.85 % (912) 427-1745 Academy Campus Athens Andraá M, Perrin 800 U.S. Highway 29 North Elbert County FULL Candidates Candidates MSN, PhD (c ), RN Technical Athens, Georgia 30601-1500 Campus 14 / 14 Pass 170 / 157 [email protected] College 100 % Pass 92.35% www.athenstech.edu (706) 213-2108 Atlanta Cassandra Danby, 1560 Metropolitan Parkway SW FULL Candidates Candidates RN, BSN Technical Atlanta, Georgia 30310-4446 2 / 2 Pass 189 / 155 Interim [email protected] College www.atlantatech.edu 100 % Pass 82.01 % (404) 225-4528 Augusta Mary Walker, MSN 3200 Augusta Tech Drive Thomson/McDuffie FULL Candidates Candidates RN Augusta, Georgia 30906 Campus 76 / 74 Pass 389 / 357 Technical [email protected] College Waynesboro / Burke www.augustatech.edu Campus 97.37 % Pass 91.77% (706)-771-4182 Georgia Board of Examiners of Licensed Practical Nurses, Approved Practical Nursing Programs Technical College Program Contact Information Other Campus Approval NCLEX-PN 5- Year 2010 Average Parent Institution PN Programs Status Director NCLEX Score 2006 - 2010 Bainbridge College Sheree Dickenson, 2500 East Shotwell Street Early County FULL Candidates Candidates www.bainbridge.edu MSN, RN, CDE Bainbridge, GA 39818-0990 Campus 69 / 57 Pass 237 / 209 sheree.dickenson@bainbridge. -

Georgia State Pledging Pool Public Depositors As of October 2015
Georgia State Pledging Pool Public Depositors as of October 2015 2009 VINEVILLE OPERATIONS ACCOUNT ATLANTA PUBLIC SCHOOLS AAHPS 401K PLAN TRUST ATLANTA REGIONAL COMMISSION ABRAHAM BALDWIN AGRICULTURAL COLLEGE ATLANTA TECHNICAL COLLEGE ADAIRSVILLE APTS LTD 1% RESERVE AU KINGSLAND DEVELOPMENT ADAIRSVILLE ELEMENTARY SCHOOL AUGUST CANAL AUTHORITY ADVANTAGE BEHAVIORAL HEALTH SYSTEMS AUGUSTA CANAL AUTH AGRI-INDUSTRIAL DEVELOPM AUGUSTA EAST CENTRAL REGIONAL HOSPITAL AIKEN TECHNICAL COLLEGE AUGUSTA PUBLIC LIBRARY AIRPORT AUTHORITY AUGUSTA RICHMOND COUNTY ALACHUA VILLAS, LTD AUGUSTA RICHMOND COUNTY COLISEUM ALBANY AREA COMMUNITY SERVICE BOARD AUGUSTA RICHMOND COUNTY COUNCIL ALBANY AREA PRIMARY HEALTH INC AUGUSTA TECHNICAL INSTITUTE ALBANY DOUGHERTY DRUG UNIT AUGUSTA-RICHMOND COUNTY COLISEUM AUTH ALBANY DOUGHERTY HOSPITAL AUTHORITY AUSTELL NATURAL GAS SYSTEM ALBANY DOUGHERTY PAYROLL DEVELOPMENT AZALEA TRACE II INC ALBANY STATE UNIVERSITY AZALEA TRACE INC ALBANY TECHNICAL COLLEGE BACON COUNTY BOARD OF HEALTH ALBANY TECHNICAL COLLEGE BAINBRIDGE DOWNTOWN DEVELOPMENT AUTH ALBANY-DOUGHERTY INNER CITY AUTHORITY BAKER COUNTY BOC ALGOOD LIMITED BAKER COUNTY BOE ALPHARETTA BUSINESS COMMUNITY BALDWIN COUNTY BOARD OF COMMISSIONERS ALPHARETTA DEVELOPMENT AUTHORITY OF BALL LIMITED LP DBA AMERICUS SUMTER COUNTY HOSPITAL AUT BANK COUNTY COMMISSIONERS APPLING COUNTY BOARD OF COMMISSIONERS BARNESVILLE ASSOCIATION LTD APPLING COUNTY BOARD OF EDUCATION BARTOW CARTERSVILLE JOINT DEV AUTH APPLING COUNTY BOARD OF HEALTH BARTOW COUNTY APPLING COUNTY DFACS BARTOW COUNTY -

To Download Introduction to Georgia's
Project Wild Teacher Resource Guide: Introduction to Georgia’s Natural History Georgia Department of Natural Resources Wildlife Resources Division Timothy S. Keyes TABLE OF CONTENTS INTRODUCTION --------------------------------------------------------------------------------- 2 Prehistoric Georgia ----------------------------------------------------------------------------- 3 Physiographic Regions ------------------------------------------------------------------------- 4 MOUNTAINS --------------------------------------------------------------------------------------- 5 CUMBERLAND PLATEAU -------------------------------------------------------------------- 6 Caves --------------------------------------------------------------------------------------------- 6 Key Plants and Animals ----------------------------------------------------------------------- 7 RIDGE AND VALLEY ---------------------------------------------------------------------------- 8 Etowah River ------------------------------------------------------------------------------------ 8 Key Plants and Animals ----------------------------------------------------------------------- -8 BLUE RIDGE --------------------------------------------------------------------------------------- 10 Cove Forests ------------------------------------------------------------------------------------- 10 Key Plants and Animals ----------------------------------------------------------------------- 11 PIEDMONT ----------------------------------------------------------------------------------------- -

Chapter 14. Wildlife and Forest Communities 341
chapteR 14. Wildlife and Forest Communities 341 Chapter 14. Wildlife and Forest communities Margaret Trani Griep and Beverly Collins1 key FindingS • Hotspot areas for plants of concern are Big Bend National Park; the Apalachicola area of the Southern Gulf Coast; • The South has 1,076 native terrestrial vertebrates: 179 Lake Wales Ridge and the area south of Lake Okeechobee amphibians, 525 birds, 176 mammals, and 196 reptiles. in Peninsular Florida; and coastal counties of North Species richness is highest in the Mid-South (856) and Carolina in the Atlantic Coastal Plain. The Appalachian- Coastal Plain (733), reflecting both the large area of these Cumberland highlands also contain plants identified by subregions and the diversity of habitats within them. States as species of concern. • The geography of species richness varies by taxa. • Species, including those of conservation concern, are Amphibians flourish in portions of the Piedmont and imperiled by habitat alteration, isolation, introduction of Appalachian-Cumberland highlands and across the Coastal invasive species, environmental pollutants, commercial Plain. Bird richness is highest along the coastal wetlands of development, human disturbance, and exploitation. the Atlantic Ocean and Gulf of Mexico, mammal richness Conditions predicted by the forecasts will magnify these is highest in the Mid-South and Appalachian-Cumberland stressors. Each species varies in its vulnerability to highlands, and reptile richness is highest across the forecasted threats, and these threats vary by subregion. Key southern portion of the region. areas of concern arise where hotspots of vulnerable species • The South has 142 terrestrial vertebrate species coincide with forecasted stressors. considered to be of conservation concern (e.g., global • There are 614 species that are presumed extirpated from conservation status rank of critically imperiled, imperiled, selected States in the South; 64 are terrestrial vertebrates or vulnerable), 77 of which are listed as threatened or and 550 are vascular plants. -

Southeast Alabama Regional Planning and Development Commission
2017-2019 Southeast Alabama Regional Planning and Development Commission Region 7 Serving the counties of: Barbour, Coffee, Covington, Dale, Geneva, Henry & Houston Prepared by the Southeast Alabama Regional Planning and Development Commission In cooperation with the Alabama Association of Regional Councils (AARC) and the Alabama Department of Transportation (ALDOT) September 2017 Reproduction of this document, in whole or in part, is permitted as long as appropriate attribution is made. The preparation of this report was financed, in part, by the Alabama Department of Transportation (ALDOT). For information or copies, contact: Southeast Alabama Regional Planning and Development Commission 462 North Oates Street P.O. Box 1406 Dothan, AL 36302 Planning Area: Barbour, Coffee, Covington, Dale, Geneva, Henry and Houston counties, including the Southeast Wiregrass Area Metropolitan Planning Organization (MPO) and the Southeast Alabama Rural Planning Organization (RPO) Lead Agency: Southeast Alabama Regional Planning & Development Commission Contact Persons: Scott Farmer, Community Development Director [email protected] Darrell Rigsby, Transportation Director [email protected] Telephone: (334) 794-4093 ii Abstract: This document is the Human Services Coordinated Transportation Plan for the Southeast Alabama Region 2017-2019, which updates the product that the Southeast Alabama Regional Planning and Development Commission (SEARP&DC) most recently published in September 2015. The Human Services Coordinated Transportation Plan for the Southeast Alabama Region 2017-2019 is developed in accordance with the Fixing America’s Surface Transportation (FAST) Act, as well as former transportation bills, including the Safe, Accountable, Flexible, Efficient Transportation Equity Act: A Legacy for Users (SAFETEA–LU) and the Moving Ahead for Progress in the 21st Century (MAP-21) federal transportation reauthorizations. -

Irrigation and Water Use Reporting in the Tennessee Valley Region of Alabama
Irrigation and Water Use Reporting in the Tennessee Valley Region of Alabama by Jarrett Roland A thesis submitted to the Graduate Faculty of Auburn University in partial fulfillment of the requirements for the Degree of Master of Science Auburn, Alabama August 3, 2019 Keywords: Irrigation, Water Resources, Sustainability, Water Use, GIS, Tennessee River Valley Copyright 2019 by Jarrett Roland Approved by: Dr. Philip Chaney, Chair, Associate Professor of Geosciences Dr. Stephanie Shepherd, Assistant Professor of Geosciences Dr. Christopher Burton, Assistant Professor of Geosciences Abstract Irrigation is the largest consumptive use of water in the United States. Accurate monitoring of water use for irrigation is critical for a state to sustainably manage its water resources. Federal programs provide valuable information for states to utilize, but individual states may develop their own program to supplement those federal programs. In 1993, Alabama passed the Water Resources Act, requiring irrigators with the capacity to withdraw more than 100,000 gallons per day to apply for a Certificate of Beneficial Use (COU) with Alabama’s Office of Water Resources for each pump that is withdrawing water and report their use annually. Recent studies have revealed issues with the effectiveness of Alabama’s COU program. This study investigated CP irrigation in the Tennessee Valley region in northern Alabama for the years 2011 to 2015 to determine whether CP irrigators were applying for COUs and reporting water use as required by the program. The study found that CP irrigation increased substantially (194 to 324; +67.0%) for the study period. However, the number of pumps registered for irrigators did not change (77) throughout the study period. -

Columbia County Growth Management Plan Update Community Assessment
Columbia County Growth Management Plan Update Community Assessment June 2005 1 / COMMUNITY ASSESSMENT COLUMBIA COUNTY GROWTH MANAGEMENT PLAN UPDATE 1.0 Population 9 Introduction .........................................................................................9 The Planning Area...............................................................................9 Population Trends .............................................................................10 Population Composition.....................................................................11 Population Projection by Age.............................................................12 Income and Educational Attainment ..................................................14 Columbia County Tomorrow..............................................................16 2.0 Housing 19 Introduction .......................................................................................19 Residential Growth in Columbia County ............................................19 Current Housing Stock ......................................................................19 Age of Housing Stock........................................................................21 Housing Growth.................................................................................21 Household Type ................................................................................22 Special Housing Needs .....................................................................23 Housing Cost.....................................................................................24