A COMPARISON of the JOINTS of the ARM and LEG and the SIGNIFICANCE of the STRUCTURAL DIFFERENCES BETWEEN THEM by C
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
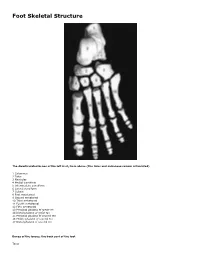
Skeletal Foot Structure
Foot Skeletal Structure The disarticulated bones of the left foot, from above (The talus and calcaneus remain articulated) 1 Calcaneus 2 Talus 3 Navicular 4 Medial cuneiform 5 Intermediate cuneiform 6 Lateral cuneiform 7 Cuboid 8 First metatarsal 9 Second metatarsal 10 Third metatarsal 11 Fourth metatarsal 12 Fifth metatarsal 13 Proximal phalanx of great toe 14 Distal phalanx of great toe 15 Proximal phalanx of second toe 16 Middle phalanx of second toe 17 Distal phalanx of second toe Bones of the tarsus, the back part of the foot Talus Calcaneus Navicular bone Cuboid bone Medial, intermediate and lateral cuneiform bones Bones of the metatarsus, the forepart of the foot First to fifth metatarsal bones (numbered from the medial side) Bones of the toes or digits Phalanges -- a proximal and a distal phalanx for the great toe; proximal, middle and distal phalanges for the second to fifth toes Sesamoid bones Two always present in the tendons of flexor hallucis brevis Origin and meaning of some terms associated with the foot Tibia: Latin for a flute or pipe; the shin bone has a fanciful resemblance to this wind instrument. Fibula: Latin for a pin or skewer; the long thin bone of the leg. Adjective fibular or peroneal, which is from the Greek for pin. Tarsus: Greek for a wicker frame; the basic framework for the back of the foot. Metatarsus: Greek for beyond the tarsus; the forepart of the foot. Talus (astragalus): Latin (Greek) for one of a set of dice; viewed from above the main part of the talus has a rather square appearance. -

Wrist Fracture – Advice Following Removal of Your Cast
Wrist Fracture – advice following removal of your cast A plaster cast usually prevents a fracture from moving, but allows your fingers to move. The cast also reduces pain. What to expect It usually takes four to six weeks for new bone to form to heal your fracture. When the cast is removed most people find that their wrist is stiff, weak and uncomfortable to start with. It may also be prone to swelling and the skin dry or flaky, this is quite normal. It is normal to get some pain after your fracture. If you need painkillers you should take them as prescribed as this will allow you to do your exercises and use your wrist for light activities. You can ask a Pharmacist about over the counter painkillers. If your pain is severe, continuous or excessive you should contact your GP. The new bone gradually matures and becomes stronger over the next few months. It is likely to be tender and may hurt if you bang it. The muscles will be weak initially, but they should gradually build up as you start to use your hand and wrist. When can I start to use my hand and wrist? It is important to try and use your hand and wrist as normally as possible. Start with light activities like fastening buttons, washing your face, eating, turning the pages of books over etc. Build up as pain allows. Avoid lifting a kettle for 4 weeks If I have been given a Wrist splint You may have been given a wrist splint to wear. -

Wrist Fracture
Hand Conditions: WRIST FRACTURE A wrist fracture is a break in one or more of the bones in the wrist. The wrist is made up of the two bones in the forearm called the radius and the ulna. It also includes eight carpal bones. The carpal bones lie between the end of the forearm bones and the bases of the fi ngers. The most commonly fractured carpal bone is called the scaphoid or navicular bone. This fact sheet will focus on fractures of the carpal bones of the wrist. Causes A wrist fracture is caused by trauma to the bones in the wrist. Trauma may be caused by: • Falling on an outstretched arm • Direct blow to the wrist • Severe twist of the wrist Risk Factors Factors that increase your chance of developing a wrist fracture include: • Participating in contact sports, such as football or soccer • Participating in activities such as in-line skating, skateboarding, or bike riding • Participating in any activity which could cause you to fall on your outstretched hand • Violence or high-velocity trauma, such as an automobile accident Symptoms If you have any of these symptoms, do not assume they are due to a wrist fracture. Symptoms of a wrist fracture include. • Pain • Swelling and tenderness around the wrist • Bruising around the wrist • Limited range of wrist or thumb motion • Visible deformity in the wrist For more information visit us online at www.ptandme.com Hand Conditions: WRIST FRACTURE Diagnosis Your doctor will ask about your symptoms, physical activity, and how the injury occurred. The injured area will be examined. -

Study Guide Medical Terminology by Thea Liza Batan About the Author
Study Guide Medical Terminology By Thea Liza Batan About the Author Thea Liza Batan earned a Master of Science in Nursing Administration in 2007 from Xavier University in Cincinnati, Ohio. She has worked as a staff nurse, nurse instructor, and level department head. She currently works as a simulation coordinator and a free- lance writer specializing in nursing and healthcare. All terms mentioned in this text that are known to be trademarks or service marks have been appropriately capitalized. Use of a term in this text shouldn’t be regarded as affecting the validity of any trademark or service mark. Copyright © 2017 by Penn Foster, Inc. All rights reserved. No part of the material protected by this copyright may be reproduced or utilized in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the copyright owner. Requests for permission to make copies of any part of the work should be mailed to Copyright Permissions, Penn Foster, 925 Oak Street, Scranton, Pennsylvania 18515. Printed in the United States of America CONTENTS INSTRUCTIONS 1 READING ASSIGNMENTS 3 LESSON 1: THE FUNDAMENTALS OF MEDICAL TERMINOLOGY 5 LESSON 2: DIAGNOSIS, INTERVENTION, AND HUMAN BODY TERMS 28 LESSON 3: MUSCULOSKELETAL, CIRCULATORY, AND RESPIRATORY SYSTEM TERMS 44 LESSON 4: DIGESTIVE, URINARY, AND REPRODUCTIVE SYSTEM TERMS 69 LESSON 5: INTEGUMENTARY, NERVOUS, AND ENDOCRINE S YSTEM TERMS 96 SELF-CHECK ANSWERS 134 © PENN FOSTER, INC. 2017 MEDICAL TERMINOLOGY PAGE III Contents INSTRUCTIONS INTRODUCTION Welcome to your course on medical terminology. You’re taking this course because you’re most likely interested in pursuing a health and science career, which entails proficiencyincommunicatingwithhealthcareprofessionalssuchasphysicians,nurses, or dentists. -

PE1897 Wrist and Hand Stretches
Patient and Family Education Wrist and Hand Stretches How can I help my child do the stretches? Use these exercises to help stretch the You play an important role in your child’s therapy. Older children may need wrist and hand. reminders to do their stretches every day. You may need to help position your younger child for the stretches. Or you may need to help stretch your child’s hand or arm. Be sure to pay attention to your child’s alignment and posture to make sure each stretch is performed correctly. How often should my child do the stretches? These stretches should be done twice a day, or as instructed by your therapist: ______________________________________________________________ Stretches Wrist extension Hold arm out in front Use opposite hand to bend wrist up with fingers straight Option to straighten elbow for increased stretch Hold for 30 seconds or _______ Repeat 2 times or ___________ VHI Wrist extension Sit with elbows on table Place palms together Slowly lower wrists to table Hold for 30 seconds or ______ Repeat 2 times or __________ VHI Wrist flexion Hold arm out in front Use opposite hand to bend wrist down Option to straighten elbow for increased stretch Option to curl fingers for increased stretch Hold for 30 seconds or ______ VHI Repeat 2 times or __________ 1 of 2 Wrist and Hand Stretches Wrist radial/ulnar deviation To Learn More Hold arm at side of body with palm • Occupational/Physical facing forward Therapy 206-987-2113 Use opposite hand to straighten wrist toward the thumb side Do not allow the wrist to flex forward to extend backward Free Interpreter Hold for 30 seconds or ______ Services Repeat 2 times or __________ • In the hospital, ask BioEx Systems Inc.* your child’s nurse. -

Standing Shoulder Flexion with Resistance
Prepared by Samantha Bohy Michigan STEP 1 STEP 2 Standing Shoulder Flexion with Resistance REPS: 15 | SETS: 2 | WEEKLY: 5x | Setup Begin in a standing upright position holding one end of a resistance band anchored under your foot with your thumb pointing forward. Movement Lift your arm straight forward to shoulder height, then slowly lower it back down and repeat. STEP 1 STEP 2 Single Arm Shoulder Extension with Resistance REPS: 15 | SETS: 2 | WEEKLY: 5x | Setup Begin standing tall, holding the end of a band that is anchored in front of you. Movement Pull your arm back, bringing your hand behind you. Return to the starting position and repeat. STEP 1 STEP 2 Standing Single Arm Shoulder Abduction with Resistance REPS: 15 | SETS: 2 | WEEKLY: 5x | Setup Begin in a standing upright position holding one end of a resistance band anchored under your feet with your thumb pointing up. Movement Lift your arm straight out to your side, to shoulder height, then lower it back down and repeat. Tip Make sure to maintain good posture and do not shrug your shoulder during the exercise. STEP 1 STEP 2 Shoulder Adduction with Anchored Resistance REPS: 15 | SETS: 2 | WEEKLY: 5x | Setup Begin in a standing upright position holding the end of a resistance band in one hand with your arm straight and palm facing downward, to the side of the anchor point. Movement Pull your arm down against the resistance band to your side, then slowly return to the starting position and repeat. STEP 1 STEP 2 Shoulder External Rotation with Anchored Resistance REPS: 15 | SETS: 2 | WEEKLY: 5x | Setup Begin standing upright with your elbow bent at 90 degrees and a towel roll tucked under your arm, holding a resistance band that is anchored out to your opposite side. -

Ulnar Nerve Injury & Repair
1/8/16 Ulnar Nerve Injury & Repair: Philadelphia’s Rehabilita6on & Ortho6c Magic Gardens Intervenon 1020 South Street Jenifer M. Haines,MS,OTR /L,CHT The Philadelphia Hand Center Outcomes Following Ulnar Nerve Repair Purpose: quan4fy variables influencing outcome aer 1. Outcomes & Expectaons ulnar & median nerve repair 2. Anatomy Methods: meta-analysis, literature review, 23 ar4cles 3. Clinical presentaon ulnar Results: nerve injury 1) 45% “sasfactory motor outcome” (71% < median nerve) 4. Func4onal deficits 41% “sasfactory sensory outcome” (approx. = median 5. Acute post-operave stage nerve) 6. ReHabilitaon stage 2) HigH level injury - poor motor outcome *irreversible motor damage by 1 ½-2 years (before re- innervaon of muscle) Ruijs, 2005 Long Term Outcomes of Ulnar 3) Paents < 16 years old 4x more likely sasfactory Nerve Injury motor recovery than those > 40 * neuroplas4city Purposes: 1. assess long term outcomes of paents following 4) Delayed surgery, lower cHance motor & sensory peripHeral repair recovery 2. determine relaonsHips between measures of Hand * Improvement possible up to 3 years func4on and nerve recovery. Methods: evaluated 32 paents approx. 5 years post-op Rosen and Lundborg scale, sensory, motor, and pain/ symptom tests, and self-report measures Ruijs, 2005 MacDermid J, 2010 1 1/8/16 Results: Paents retained 82.91% global func4on: 44.48%-84.90% sensory funcon “Factors that predict outcomes aer the repair of 80.13%-89.89% motor func4on peripHeral nerve injuries of the upper limb include age, 89.75%-93.19% pain/symptom experience gender, repair 4me, repair materials, defect length, duraon of follow-up and nerve injured.” Self-report measures of Hand func4on were more closely related to nerve recovery than were pHysical measures. -

Nerve Blocks for Surgery on the Shoulder, Arm Or Hand
The Association of Regional The Royal College of Anaesthetists of Great Anaesthesia – Anaesthetists Britain and Ireland United Kingdom Nerve blocks for surgery on the shoulder, arm or hand Information for patients and families www.rcoa.ac.uk/patientinfo First edition 2015 This leaflet is for anyone who is thinking about having a nerve block for an operation on the shoulder, arm or hand. It will be of particular interest to people who would prefer not to have a general anaesthetic. The leaflet has been written with the help of patients who have had a nerve block for their operation. You can find more information leaflets on the website www.rcoa.ac.uk/patientinfo. The leaflets may also be available from the anaesthetic department or pre-assessment clinic in your hospital. The website includes the following: ■ Anaesthesia explained (a more detailed booklet). ■ You and your anaesthetic (a shorter summary). ■ Your spinal anaesthetic. ■ Anaesthetic choices for hip or knee replacement. ■ Epidural pain relief after surgery. ■ Local anaesthesia for your eye operation. ■ Your child’s general anaesthetic. ■ Your anaesthetic for major surgery with planned high dependency care afterwards. ■ Your anaesthetic for a broken hip. Risks associated with your anaesthetic This is a collection of 14 articles about specific risks associated with having an anaesthetic or an anaesthetic procedure. It supplements the patient information leaflets listed above and is available on the website: www.rcoa.ac.uk/patients-and-relatives/risks. Throughout this leaflet and others in the series, we have used this symbol to highlight key facts. 2 NERVE BLOCKS FOR SURGERY ON THE SHOULDER, ARM OR HAND Brachial plexus block? The brachial plexus is the group of nerves that lies between your neck and your armpit. -

Canine Tarsus Stabilization Surgical Technique 1 2
Canine Tarsus Stabilization Surgical Technique 1 2 Place the patient in a dorsal recumbent position and Make a medial incision beginning just proximal to the administer general anesthesia. Perform a hanging limb medial malleolus extending distally to the level of the technique with aseptic preparation and appropriate proximal intertarsal joint. Inspect the medial structures limb draping. of the tarsal joint. Identify the origin of the long part of the medial collateral ligament (MCL) on the medial malleolus of the tibia. 3 4 Using the aiming guide, place the 0.049 inch (1.2 mm) Use the 2.0 mm cannulated drill over the guidewire to guidewire from the origin of the long part of the MCL create the tibial bone tunnel for suture passage. in a proximal and slight cranial direction such that the it emerges on the lateral tibia just cranial to the fibula. Make a small incision on the lateral side where the guidewire tents the skin. 5 As a surgical landmark, identify the talocentral tarsal joint on the medial side by either a palpation or by inserting a small needle into the joint space. Using the aiming guide, place a 0.049 inch (1.2 mm) guidewire starting at the medial aspect of the talar head oriented parallel to the talocentral tarsal joint in a slightly plantar direction to capture the calcaneus. The guidewire should emerge just plantar to the boney tubercle of the lateral portion of the distal calcaneus. Make a small incision where the guidewire tents the skin. 6 7 Use the 2.0 mm cannulated drill over the guidewire to Identify the insertion of the short part of the MCL on the create the distal talus bone tunnel for suture passage. -

Locking Small Fragment Overview
Surgical Technique Locking Small Fragment Overview PERI-LOC™ Locked Plating System Locking Small Fragment Overview Surgical Technique Table of contents Product overview ...................................................................................2 Introduction .............................................................................................2 Indications ...............................................................................................2 Design features and benefits .................................................................3 System overview ....................................................................................4 Implant overview ...................................................................................10 Surgical technique ................................................................................23 Fracture reduction ...................................................................................23 Clavicle Locking Plate .............................................................................24 3.5mm Proximal Humerus Locking Plate ................................................25 Distal Humerus Locking Plates ...............................................................26 Olecranon Locking Plate .........................................................................28 3.5mm Lateral Proximal Tibia Locking Plate ......................................... 29 3.5mm Medial Distal Tibia Locking Plate .............................................. 30 3.5mm Anterolateral -

Bone Limb Upper
Shoulder Pectoral girdle (shoulder girdle) Scapula Acromioclavicular joint proximal end of Humerus Clavicle Sternoclavicular joint Bone: Upper limb - 1 Scapula Coracoid proc. 3 angles Superior Inferior Lateral 3 borders Lateral angle Medial Lateral Superior 2 surfaces 3 processes Posterior view: Acromion Right Scapula Spine Coracoid Bone: Upper limb - 2 Scapula 2 surfaces: Costal (Anterior), Posterior Posterior view: Costal (Anterior) view: Right Scapula Right Scapula Bone: Upper limb - 3 Scapula Glenoid cavity: Glenohumeral joint Lateral view: Infraglenoid tubercle Right Scapula Supraglenoid tubercle posterior anterior Bone: Upper limb - 4 Scapula Supraglenoid tubercle: long head of biceps Anterior view: brachii Right Scapula Bone: Upper limb - 5 Scapula Infraglenoid tubercle: long head of triceps brachii Anterior view: Right Scapula (with biceps brachii removed) Bone: Upper limb - 6 Posterior surface of Scapula, Right Acromion; Spine; Spinoglenoid notch Suprspinatous fossa, Infraspinatous fossa Bone: Upper limb - 7 Costal (Anterior) surface of Scapula, Right Subscapular fossa: Shallow concave surface for subscapularis Bone: Upper limb - 8 Superior border Coracoid process Suprascapular notch Suprascapular nerve Posterior view: Right Scapula Bone: Upper limb - 9 Acromial Clavicle end Sternal end S-shaped Acromial end: smaller, oval facet Sternal end: larger,quadrangular facet, with manubrium, 1st rib Conoid tubercle Trapezoid line Right Clavicle Bone: Upper limb - 10 Clavicle Conoid tubercle: inferior -
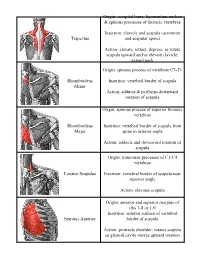
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: