The Gully a Scientific Review of Its Environment and Ecosystem
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Characteristics and Importance of Rill and Gully Erosion: a Case Study in a Small Catchment of a Marginal Olive Grove
Cuadernos de Investigación Geográfica 2015 Nº 41 (1) pp. 107-126 ISSN 0211-6820 DOI: 10.18172/cig.2644 © Universidad de La Rioja CHARACTERISTICS AND IMPORTANCE OF RILL AND GULLY EROSION: A CASE STUDY IN A SMALL CATCHMENT OF A MARGINAL OLIVE GROVE E.V. TAGUAS1*, E. GUZMÁN1, G. GUZMÁN1, T. VANWALLEGHEM1, J.A. GÓMEZ2 1Rural Engineering Department/ Agronomy Department, University of Cordoba, Campus Rabanales, Leonardo Da Vinci building, 14071 Córdoba, Spain. 2Institute for Sustainable Agriculture, CSIC, Apartado 4084, 14080 Córdoba, Spain. ABSTRACT. Measurements of gullies and rills were carried out in an olive or- chard microcatchment of 6.1 ha over a 4-year period (2010-2013). No tillage management allowing the development of a spontaneous grass cover was imple- mented in the study period. Rainfall, runoff and sediment load were measured at the catchment outlet. The objectives of this study were: 1) to quantify erosion by concentrated flow in the catchment by analysis of the geometric and geomor- phologic changes of the gullies and rills between July 2010 and July 2013; 2) to evaluate the relative percentage of erosion derived from concentrated runoff to total sediment yield; 3) to explain the dynamics of gully and rill formation based on the hydrological patterns observed during the study period; and 4) to improve the management strategies in the olive grove. Control sections in gullies were established in order to get periodic measurements of width, depth and shape in each campaign. This allowed volume changes in the concentrated flow network to be evaluated over 3 periods (period 1 = 2010-2011; period 2 = 2011-2012; and period 3 = 2012-2013). -

Alluvial Fans in the Death Valley Region California and Nevada
Alluvial Fans in the Death Valley Region California and Nevada GEOLOGICAL SURVEY PROFESSIONAL PAPER 466 Alluvial Fans in the Death Valley Region California and Nevada By CHARLES S. DENNY GEOLOGICAL SURVEY PROFESSIONAL PAPER 466 A survey and interpretation of some aspects of desert geomorphology UNITED STATES GOVERNMENT PRINTING OFFICE, WASHINGTON : 1965 UNITED STATES DEPARTMENT OF THE INTERIOR STEWART L. UDALL, Secretary GEOLOGICAL SURVEY Thomas B. Nolan, Director The U.S. Geological Survey Library has cataloged this publications as follows: Denny, Charles Storrow, 1911- Alluvial fans in the Death Valley region, California and Nevada. Washington, U.S. Govt. Print. Off., 1964. iv, 61 p. illus., maps (5 fold. col. in pocket) diagrs., profiles, tables. 30 cm. (U.S. Geological Survey. Professional Paper 466) Bibliography: p. 59. 1. Physical geography California Death Valley region. 2. Physi cal geography Nevada Death Valley region. 3. Sedimentation and deposition. 4. Alluvium. I. Title. II. Title: Death Valley region. (Series) For sale by the Superintendent of Documents, U.S. Government Printing Office Washington, D.C., 20402 CONTENTS Page Page Abstract.. _ ________________ 1 Shadow Mountain fan Continued Introduction. ______________ 2 Origin of the Shadow Mountain fan. 21 Method of study________ 2 Fan east of Alkali Flat- ___-__---.__-_- 25 Definitions and symbols. 6 Fans surrounding hills near Devils Hole_ 25 Geography _________________ 6 Bat Mountain fan___-____-___--___-__ 25 Shadow Mountain fan..______ 7 Fans east of Greenwater Range___ ______ 30 Geology.______________ 9 Fans in Greenwater Valley..-----_____. 32 Death Valley fans.__________--___-__- 32 Geomorpholo gy ______ 9 Characteristics of fans.._______-___-__- 38 Modern washes____. -

SCI Lecture Paper Series
SCI LECTURE PAPERS SERIES HIGHWAY DRAINAGE SYSTEMS Santi V Santhalingam Highways Agency, Room 4/41, St. Christopher House Southwark Street, London SE1 0TE Telephone +44 (0) 171 921 4954 Fax +44 (0) 171 921 4411 © Highways Agency 1999 copyright reserved ISSN 1353-114X LPS 102/99 Key words highways, drainage, runoff, surface, sub-surface, systems INTRODUCTION Appropriate drainage is an important feature of good highway design in terms of ensuring required level of service and value for money are achieved. Highway drainage has two major objectives: safety of the road user and longevity of the pavement. Speedy removal of surface water will help to ensure safe and comfortable conditions for the road user. Provision of effective sub-drainage will maximise longevity of the pavement and its associated earthworks. Highway drainage can therefore be broadly classified into two elements – surface run-off and sub-surface run-off: these two elements are not completely disparate in that some of the surface water may find its way into the road foundation through surfaces which are not completely impermeable thence requiring removal by sub-drainage. Based on these fundamental principles, drainage methods in the UK are broadly divided into two categories: (a) combined systems, where the surface and sub-surface water are collected and transported in the same pipe, and (b) separate systems, where the two elements are collected and transported in separate pipes Within the broader definition of the two systems there are a number of different drainage methods that are in use on UK highways, some of them more common than others. -
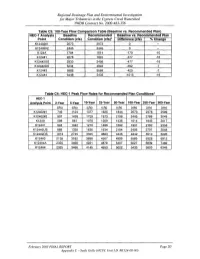
Regional Drainage Plan and Environmental Investigation for Major Tributaries in the Cypress Creek Watershed TWDB Contract No
Regional Drainage Plan and Environmental Investigation for Major Tributaries in the Cypress Creek Watershed TWDB Contract No. 2000-483-356 Table C3: 100-Year Flow Comparison Table (Baseline vs. Recommended Plan) HEC-1 Analysis Baseline Recommended Baseline vs. Recommended Plan Point Condition (cfs) Condition (cfs)· Difference (cfs) "10 Change K12402#1 2073 2073 0 -- K12402#2 2445 2445 0 -- K124A 1784 1614 -170 -10 K124#1 2278 1901 -377 -16 K124#2US 2933 2456 -477 -16 K124#2DS 5234 4842 -392 -7 K124#3 5989 5569 -420 -7 K124#4 6448 5433 -1015 -16 Table C4' HEC-1 Peak Flow Rates for Recommended Plan Conditions· HEC-1 Analysis Point 2-Year S-Year 10-Year 2S-Year SO-Year 100-Year 2S0-Year SOO-Year (cis) (cis) (cis) (cis) (cis) (cis) (cis) ~cls) K12402#1 746 1124 1377 1625 1844 2073 2376 2599 K12402#2 937 1428 1729 1973 2199 2445 2788 3049 K124A 588 881 1076 1269 1436 1614 1845 2017 K124#1 694 1042 1270 1496 1692 1901 2162 2358 K124#2US 889 1339 1636 1934 2184 2456 2797 3048 K124#2DS 1813 2745 3355 3883 4346 4842 5510 6026 K124#3 2136 3182 3898 4507 4999 5569 6328 6912 K124#4A 2325 3466 4221 4878 5407 6027 6839 7462 K124#4 2325 3466 4145 4653 5022 5433 5953 6349 February 2003 FINAL REPORT Page 20 Appendix C - Seals Gully (HCFC Unit I.D. #KI24-00-00) Regional Drainage Plan and Environmental Investigation for Major Tributaries in the Cypress Creek Watershed TWDB Contract No. 2000-483-356 Table C5: Comparison of Water Surface Elevations (100-Year) Seals Gully (K124-00-00' Baseline Condition Recommended Plan Difference Station Location Flow WSEL -

The Gully Marine Protected Area Data Assessment
Canadian Science Advisory Secretariat (CSAS) Research Document 2015/056 Maritimes Region The Gully Marine Protected Area Data Assessment K. Allard1, N. Cochrane2, K. Curran2, D. Fenton2, T. Koropatnick2, C. Gjerdrum3, B.J.W. Greenan2, E. Head2, P. Macnab2, H. Moors-Murphy2, A. Serdynska2, M.K. Trzcinski2, M. Vaughan4, and H. Whitehead5 Edited by A. Moore2 1 Canadian Wildlife Service Environment Canada 17 Waterfowl Lane, Sackville, NB E4L 1G6 2 Bedford Institute of Oceanography Fisheries and Oceans Canada P.O. Box 1006, Dartmouth NS B2Y 4A2 3 Canadian Wildlife Service Environment Canada 45 Alderney Drive, Dartmouth, NS B2Y 2N6 4 Department of Zoology University of British Columbia 6270 University Blvd., BC V6T 1Z4 5 Department of Biology Dalhousie University 1355 Oxford Street, Halifax, NS B3H 4J1 December 2015 Foreword This series documents the scientific basis for the evaluation of aquatic resources and ecosystems in Canada. As such, it addresses the issues of the day in the time frames required and the documents it contains are not intended as definitive statements on the subjects addressed but rather as progress reports on ongoing investigations. Research documents are produced in the official language in which they are provided to the Secretariat. Published by: Fisheries and Oceans Canada Canadian Science Advisory Secretariat 200 Kent Street Ottawa ON K1A 0E6 http://www.dfo-mpo.gc.ca/csas-sccs/ [email protected] © Her Majesty the Queen in Right of Canada, 2015 ISSN 1919-5044 Correct citation for this publication: Allard, K., Cochrane, N., Curran, K., Fenton, D., Koropatnick, T., Gjerdrum, C., Greenan, B.J.W., Head, E., Macnab, P., Moors-Murphy, H., Serdynska, A., Trzcinski, M.K., Vaughan, M., and Whitehead, H. -

Technical Supplement 14P--Gullies and Their Control
Technical Gullies and Their Control Supplement 14P (210–VI–NEH, August 2007) Technical Supplement 14P Gullies and Their Control Part 654 National Engineering Handbook Issued August 2007 Cover photo: Gully erosion may be a significant source of sediment to the stream. Gullies may also form in the streambanks due to uncontrolled flows from the flood plain (valley trenches). Advisory Note Techniques and approaches contained in this handbook are not all-inclusive, nor universally applicable. Designing stream restorations requires appropriate training and experience, especially to identify conditions where various approaches, tools, and techniques are most applicable, as well as their limitations for design. Note also that prod- uct names are included only to show type and availability and do not constitute endorsement for their specific use. (210–VI–NEH, August 2007) Technical Gullies and Their Control Supplement 14P Contents Purpose TS14P–1 Introduction TS14P–1 Classical gullies and ephemeral gullies TS14P–3 Gullying processes in streams TS14P–3 Issues contributing to gully formation or enlargement TS14P–5 Land use practices .........................................................................................TS14P–5 Soil properties ................................................................................................TS14P–6 Climate ............................................................................................................TS14P–6 Hydrologic and hydraulic controls ..............................................................TS14P–6 -

Gully Erosion and Its Causes
Gully Erosion – Part 1 GULLY EROSION AND ITS CAUSES Gully erosion – “A complex of processes whereby the removal of soil is characterised by incised channels in the landscape.” NSW Soil Conservation Service, 1986. Photo 1 – Head of gully erosion (Qld) Photo 2 – Rural gully erosion (SA) Introduction Gully erosion is usually distinguished from the boarder term ‘watercourse erosion’ by the path the erosion follows, which is normally along an overland flow path rather than along a creek. Prior to the occurrence of the gully erosion, the overland flow path would likely have carried only shallow concentrated flows conveying stormwater runoff to a down-slope watercourse. Gully erosion, however, can also occur within a watercourse, typically within the upper reaches, and typically resulting from an active ‘head-cut’ migrating rapidly up the valley. Gully erosion is best characterised as a ‘bed instability’ that subsequently causes in ‘bank instabilities’. Unstable banks maybe the most visible aspect of gully erosion; but stabilisation of a gully normally needs to start with stabilisation of the gully bed. The mechanics of gully erosion Most gullies start out as shallow overland flow paths that carry flows only during periods of heavy rainfall (Figure 1). An extraordinary event of some type causes an initial erosion point or ‘nick’ point (Photo 3) to form somewhere along the drainage path. A bell-shaped scour hole sometimes forms at the head of the gully (Photo 4), which is usually deeper than the immediate downstream gully bed. Photo 3 – ‘Nick’ point representing the Photo 4 – Early stage of gully erosion early stages of gully erosion (SA) within an urban overland flow path (Qld) © Catchments & Creeks Pty Ltd April 2010 Page 1 The initial nick point usually occurs at the downstream end of the gully, and usually at a significant change in grade along the flow path, such as the point where the overland flow spills into a watercourse. -

Gum Gully (Unclassified Water Body) Segment: 0508B Sabine River Basin
2002 Texas Water Quality Inventory Page : 1 (based on data from 03/01/1996 to 02/28/2001) Gum Gully (unclassified water body) Segment: 0508B Sabine River Basin Basin number: 5 Basin group: A Water body description: From the confluence of Adams Bayou to the upstream perennial portion of the stream northwest of Orange in Orange County Water body classification: Unclassified Water body type: Freshwater Stream Water body length / area: 3.5 Miles Water body uses: Aquatic Life Use, Contact Recreation Use, Fish Consumption Use Standards Not Met and Concerns in Previous Years Support Status Assessment Area Use or Concern Parameter Category Entire creek Aquatic Life Use Not Supporting depressed dissolved oxygen 5c Entire creek Contact Recreation Use Not Supporting bacteria 5c Additional Information: The fish consumption use was not assessed. This water body was identified on the 2000 303(d) List as not supporting the contact recreation use due to bacteria. Because there were insufficient data available in 2002 to evaluate changes in water quality, this water body will be identified as not meeting the standard for bacteria until sufficient data are available to demonstrate use support. This water body was also identified on the 2000 303(d) List as not supporting the aquatic life use due to depressed dissolved oxygen. Because an insufficient number of 24-hour dissolved oxygen values were available in 2002 to determine if the criterion is supported, this water body will be identified as not meeting the standard for dissolved oxygen until sufficient 24-hour measurements are available to demonstrate support of the criterion. -

Rill and Gully Formation Following the 2010 Schultz Fire
Rill and Gully Formation Following the 2010 Schultz Fire Daniel G. Nearya, Karen A. Koestnera, Ann Youbergb, Peter E. Koestnerc, aUSDA Forest Service, Rocky Mountain Research Station, 2500 Pine Knoll Drive, Flagstaff, Arizona 86001 bArizona Geological Survey, 416 Congress Street, Suite 100, Tucson, Arizona 85701 cUSDA Forest Service, Rocky Mountain Research Station, Tonto National Forest, 2324 East McDowell Road, Phoenix, AZ 85006 (USA). Abstract: The Schultz Fire burned 6,100 ha on the eastern slopes of the San Francisco Peaks across moderate to very steep ponderosa pine and mixed conifer watersheds. There was widespread occurrence of high severity fire, with several watersheds classified as over 50% high severity. This resulted in moderate to severe water repellency in most soils, especially those on steep slopes. Prior to the fire there were no rills or gullies as the soil was protected by a thick O horizon. This protective organic layer was consumed leaving the soil exposed to raindrop impact and erosion. A series of flood events beginning in mid-July 2010 initiated erosion from the upper slopes of the watersheds. Substantial amounts of soil was eroded from hillslopes via a newly formed rill and gully system, removing the A horizon and much of the B horizon. The development of an extensive rill and gully network fundamentally changed the hydrologic response of the upper portions of every catchment. The intense, short duration rainfall of the 2010 monsoon interacted with slope, water repellency and extensive areas of bare soil to produce flood flows an order of magnitude in excess of flows produced by similar pre-fire rainfall events. -

Gully Treatment
_________________________________________________________________________________________________________ United States Part 650 Department of Engineering Field Handbook Agriculture Natural Resources Conservation Service Chapter 10 (650.10) Gully Treatment 1 ENGINEERING FIELD HANDBOOK (650‐EFH) Chapter 10 (650.10) – Gully Treatment Acknowledgments This major chapter revision was prepared under the general direction of Wayne Bogovich, PE, national agricultural engineer, Natural Resources Conservation Service (NRCS), Washington, DC, with assistance from Tony G. Funderburk, PE, agricultural engineer, NRCS, Central National Technology Support Center, Fort Worth, Texas. Extensive comments and edits were supplied by Jon Fripp, PE, stream mechanics engineer, NRCS, Fort Worth, Texas and Kerry Robinson, Ph.D., PE, hydraulic engineer, NRCS, Greensboro, North Carolina. September 2010 2 Table of Contents 1. GENERAL ........................................................................................................................................... 4 DEFINITION ................................................................................................................................... 4 INTRODUCTION ........................................................................................................................... 4 CAUSES ......................................................................................................................................... 4 2. PLANNING ......................................................................................................................................... -

Modelling Coastal Processes at Shippagan Gully Inlet, New Brunswick, Canada
4th Specialty Conference on Coastal, Estuary and Offshore Engineering 4e Conférence spécialisée sur l’ingénierie côtière et en milieu maritime Montréal, Québec May 29 to June 1, 2013 / 29 mai au 1 juin 2013 MODELLING COASTAL PROCESSES AT SHIPPAGAN GULLY INLET, NEW BRUNSWICK, CANADA A. Cornett1, M. Provan2, I. Nistor3, A. Drouin4 1 Leader, Marine Infrastructure Program, National Research Council, Ottawa, Canada 2 Graduate Student, Dept. of Civil Engineering, University of Ottawa, Canada 3 Associate Professor, Dept. of Civil Engineering, University of Ottawa, Canada 4 Senior Engineer, Public Works and Government Services Canada, Quebec, Canada Abstract: This paper describes the development, calibration and application of a numerical model of the hydrodynamic and sedimentary processes at a dynamic tidal inlet known as Shippagan Gully, located on the Gulf of St-Lawrence near Le Goulet, New Brunswick. The new model has been developed to provide guidance concerning the response of the inlet mouth to various potential interventions aimed at increasing navigation safety. The new model is based on coupling the most recent CMS-Flow and CMS- Wave models developed by the US Army Corps of Engineers. The coupled model is capable of simulating the depth-averaged currents generated within Shippagan Gully and along the neighbouring coastline due to the effects of tides, winds and waves; the transport of non-cohesive sediments; and the resulting changes in seabed morphology. The development of the model and the steps taken to calibrate and validate it against field measurements are described. The application of the model to predict the coastal processes and the response of the inlet mouth to several storms is described and discussed. -

State of the Eastern Scotian Shelf Ecosystem
Maritimes Region Ecosystem Status Report 2003/004 State of the Eastern Scotian Shelf Ecosystem Background The Eastern Scotian Shelf, comprising NAFO Div. 4VW, is a large geographic area (~108,000 km 2) supporting a wide range of ocean uses such as fisheries, oil and gas exploration and development, and shipping. It is currently the focus for the development of an integrated management plan intended to harmonize the conduct of the various ocean use activities within it (referred to as Eastern Scotian Shelf Integrated Management or ESSIM). The area is unique for having a year-round closure for directed fishing of groundfish since 1987, associated Summary with Emerald and Western Banks. In addition, The Many features of the Eastern Scotian Shelf Gully has been declared a pilot marine protected ecosystem have changed dramatically area. during the past thirty years: The Eastern Scotian Shelf consists of a series of • A major cooling event of the bottom outer shallow banks and inner basins separated by waters occurred in the mid-1980s that gullies and channels. The mean surface circulation is persisted for a decade and recent dominated by southwestward flow, much of which intensive stratification in the surface originates from the Gulf of St. Lawrence with anticyclonic circulation tending to occur over the layer has been apparent; both banks and cyclonic circulation around the basins. The phenomena are associated with flow northeastern region of the Shelf is the southern- most from upstream areas. limit of winter sea ice in the Atlantic Ocean. • The index of zooplankton abundance This document provides an assessment of the current was low in the decade of the 1990s state of the Eastern Scotian Shelf ecosystem.