Disentangling the Effects of Disturbance and Habitat Size on Stream Community Structure
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Upper Riccarton Cemetery 2007 1
St Peter’s, Upper Riccarton, is the graveyard of owners and trainers of the great horses of the racing and trotting worlds. People buried here have been in charge of horses which have won the A. J. C. Derby, the V.R.C. Derby, the Oaks, Melbourne Cup, Cox Plate, Auckland Cup (both codes), New Zealand Cup (both codes) and Wellington Cup. Area 1 Row A Robert John Witty. Robert John Witty (‘Peter’ to his friends) was born in Nelson in 1913 and attended Christchurch Boys’ High School, College House and Canterbury College. Ordained priest in 1940, he was Vicar of New Brighton, St. Luke’s and Lyttelton. He reached the position of Archdeacon. Director of the British Sailors’ Society from 1945 till his death, he was, in 1976, awarded the Queen’s Service Medal for his work with seamen. Unofficial exorcist of the Anglican Diocese of Christchurch, Witty did not look for customers; rather they found him. He said of one Catholic lady: “Her priest put her on to me; they have a habit of doing that”. Problems included poltergeists, shuffling sounds, knockings, tapping, steps tramping up and down stairways and corridors, pictures turning to face the wall, cold patches of air and draughts. Witty heard the ringing of Victorian bells - which no longer existed - in the hallway of St. Luke’s vicarage. He thought that the bells were rung by the shade of the Rev. Arthur Lingard who came home to die at the vicarage then occupied by his parents, Eleanor and Archdeacon Edward Atherton Lingard. In fact, Arthur was moved to Miss Stronach’s private hospital where he died on 23 December 1899. -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

“Glacial Lake Speight”, New Zealand? an Example for the Validity of Detailed Geomorphological Assessment with the Study of Mountain Glaciations
Express report E&G Quaternary Sci. J., 67, 25–31, 2018 https://doi.org/10.5194/egqsj-67-25-2018 © Author(s) 2018. This work is distributed under the Creative Commons Attribution 4.0 License. Disestablishing “Glacial Lake Speight”, New Zealand? An example for the validity of detailed geomorphological assessment with the study of mountain glaciations Stefan Winkler1, David Bell2, Maree Hemmingsen3, Kate Pedley2, and Anna Schoch4 1Department of Geography and Geology, University of Würzburg, Am Hubland, 97074 Würzburg, Germany 2Department of Geological Sciences, University of Canterbury, Private Bag 4800, Christchurch 8140, New Zealand 3Primary Science Solutions Ltd., Woodbury Street 75, Russley, Christchurch 8042, New Zealand 4Department of Geography, University of Bonn, Meckenheimer Allee 166, 53115 Bonn, Germany Correspondence: Stefan Winkler ([email protected]) Relevant dates: Received: 30 May 2018 – Revised: 10 August 2018 – Accepted: 21 August 2018 – Published: 28 August 2018 How to cite: Winkler, S., Bell, D., Hemmingsen, M., Pedley, K., and Schoch, A.: Disestablishing “Glacial Lake Speight”, New Zealand? An example for the validity of detailed geomorphological assessment with the study of mountain glaciations, E&G Quaternary Sci. J., 67, 25–31, https://doi.org/10.5194/egqsj- 67-25-2018, 2018. 1 Introduction implications beyond these fluvial aspects. Palaeoseismolog- ical studies claim to have detected signals of major Alpine The middle Waimakariri River catchment in the Southern Fault earthquakes in coastal environments along the eastern Alps of New Zealand, informally defined here as its reach up- seaboard of the South Island (McFadgen and Goff, 2005). stream of Waimakariri Gorge to the junction of Bealey River This requires high connectivity between the lower reaches of (Fig. -
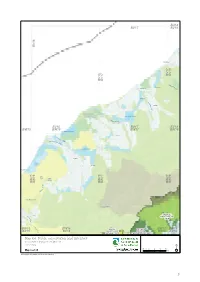
Draft Canterbury CMS 2013 Vol II: Maps
BU18 BV17 BV18 BV16 Donoghues BV17 BV18 BV16 BV17 M ik onu Fergusons i R iv Kakapotahi er Pukekura W a i ta h Waitaha a a R iv e r Lake Ianthe/Matahi W an g anui Rive r BV16 BV17 BV18 BW15 BW16 BW17 BW18 Saltwater Lagoon Herepo W ha ta ro a Ri aitangi ver W taon a R ive r Lake Rotokino Rotokino Ōkārito Lagoon Te Taho Ōkārito The Forks Lake Wahapo BW15 BW16 BW16 BW17 BW17 BW18 r e v i R to ri kā Ō Lake Mapourika Perth River Tatare HAKATERE W ai CONSERVATION h o R PARK i v e r C a l le r y BW15 R BW16 AORAKI TE KAHUI BW17 BW18 iv BX15 e BX16 MOUNT COOK KAUPEKA BX17 BX18 r NATIONAL PARK CONSERVATION PARK Map 6.6 Public conservation land inventory Conservation Management Strategy Canterbury 01 2 4 6 8 Map 6 of 24 Km Conservation unit data is current as of 21/12/2012 51 Public conservation land inventory Canterbury Map table 6.7 Conservation Conservation Unit Name Legal Status Conservation Legal Description Description Unit number Unit Area I35028 Adams Wilderness Area CAWL 7143.0 Wilderness Area - s.20 Conservation Act 1987 - J35001 Rangitata/Rakaia Head Waters Conservation Area CAST 53959.6 Stewardship Area - s.25 Conservation Act 1987 Priority ecosystem J35002 Rakaia Forest Conservation Area CAST 4891.6 Stewardship Area - s.25 Conservation Act 1987 Priority ecosystem J35007 Marginal Strip - Double Hill CAMSM 19.8 Moveable Marginal Strip - s.24(1) & (2) Conservation Act 1987 - J35009 Local Purpose Reserve Public Utility Lake Stream RALP 0.5 Local Purpose Reserve - s.23 Reserves Act 1977 - K34001 Central Southern Alps Wilberforce Conservation -

2017 Salmon Report Draft
2019 SALMON MONITORING REPORT Steve Terry August 2019 INTRODUCTION North Canterbury Fish & Game Council has been monitoring sea-run chinook salmon returns for 27 years. The South Island’s East Coast salmon fishery has been steadily declining over the last decade, with very low returns to all rivers in 2018 and only marginally improved returns in 2019. At present, fishery managers only have a limited number of options to try and ensure adequate salmon numbers reach their spawning grounds each year, the key tool being tighter regulations to reduce harvest. There is an acceptance by both North Canterbury (NC) and Central South Island Fish & Game (CSI) councils that we need to significantly reduce the harvest of wild salmon, in order to increase the numbers of fish returning to the spawning streams and rebuild the fishery. While we do not know the minimum number of spawning salmon required to sustain the population in each spawning stream or catchment, we do know that in the last decade salmon returns have steadily declined to record low levels. Regulations to incrementally reduce harvest have been put in place for the 2019/20 season, however introduction of a seasonal catch limit has been recommended by scientists as the least harmful regulation to reduce harvest and rebuild spawning numbers. It is possible that life history, genetic diversity and population resilience may be adversely affected by shortening the season and areas anglers can fish (R. Gabrielsson pers. comm). While fishery scientists do not believe the salmon fishery is at risk of extinction, there is growing concern that the increasing proportion of the run caught by anglers during a period of decreasing run size may hinder recovery when conditions eventually favour their survival at sea. -

Arthur's Pass National Park Management Plan
Arthur’s Pass National Park Management Plan Ka ü ki mata Nuku Ka ü ki mata Rangi Ka ü ki tënei whenua Hei whenua, hei kai mau te ate o te tauhou Hold fast to the land Hold fast to the sky Hold fast to this land Lest it may be treasured by others in time “A sense of history I find it consistent with a sense of history to look forward as well as backward. I study the future as much in contrast to the past as in terms of it. What will the Waimakariri Valley hold for young mountaineers in the year 1999? Will it be so full of heliports or autobahns that even the sandflies will feel themselves to be displaced insects?” Pascoe, J. 1965 Arthur's Pass National Park Management Plan Published by Department of Conservation Te Papa Atawhai Canterbury Conservancy Private Bag 4715 Christchurch December 2007. Cover: William leads the way on the Bealey Valley track through a clearing in mountain beech forest, being ‘watched over’ by a Mäori traveller (with thanks to Geoffrey Cox for the art-work); Rome and Goldney Ridges converging in the background on Mount Rolleston Kaimatau ISBN 978-0-478-14275-4 (hard copy) ISBN 978-0-478-14276-1 (CD) ISBN 978-0-478-14277-8 (Web pdf) ISSN-1171-5391-14 Canterbury Conservancy Management Planning Series No. 14 Arthur’s Pass National Park Management Plan 2007 2 CONTENTS Preface 7 How to use this plan 9 Administration of the Park 9 1 Introduction 1.1 Management Planning 11 1.2 Legislative Context 1.2.1 The National Parks Act 1980 12 1.2.1.1 National Park Bylaws 1981 12 1.2.2 The General Policy for National Parks 2005 13 -

The Christchurch Tramper
TTHEHE CCHRISTCHURCHHRISTCHURCH TRAMPERRAMPER Published by CHRISTCHURCHT TRAMPING CLUB INC PO Box 527, Christchurch. www.ctc.org.nz Affiliated with the Federated Mountain Clubs of NZ Inc. Any similarity between the opinions expressed in this newsletter and Club policy is purely coincidental. Vol. 84 December 2014/January 2015 No. 8 The CHRISTCHURCH TRAMPING CLUB has members of all ages, and runs tramping trips every weekend, ranging from easy (minimal experience required) to hard (high fitness and experience required). We also organise instructional courses and hold weekly social meetings. We have a club hut in Arthurs Pass and have gear available for hire to members. Membership rates per year are $45 member, $65 couple, $25 junior or associate, with a $5 discount for members who opt to obtain this newsletter electronically. A Frozen Lake Angelus For more about how the club operates, see the last two pages. News CHANGE OF CLUB ROOM VENUE: CHANGE OF CLUB ROOM VENUE to University of Canterbury, Room 533, Rutherford Building, effective 1 FEB 2015. We are leaving the Canterbury Horticultural Centre and moving the club room to the University of Canterbury for 2015 from 1 Feb 2015. Security doors will be open for entry on Wednesday evenings from 7:30pm to 9:30pm. See the club website (About the CTC : Where do we meet?) for a map showing the location of the room. Obituaries Doug Airey: Doug Airey, who died on the 11 November this year, was a long-standing CTC member. He joined back in 1962, was club treasurer in the early 1960s, and was still an Associate Member 52 years later until his resignation this year. -

Supplement of Disestablishing “Glacial Lake Speight”
Supplement of E&G Quaternary Sci. J., 67, 25–31, 2018 https://doi.org/10.5194/egqsj-67-25-2018-supplement © Author(s) 2018. This work is distributed under the Creative Commons Attribution 4.0 License. Supplement of Disestablishing “Glacial Lake Speight”, New Zealand? An example for the validity of detailed geomorphological assessment with the study of mountain glaciations Stefan Winkler et al. Correspondence to: Stefan Winkler ([email protected]) The copyright of individual parts of the supplement might differ from the CC BY 4.0 License. Supporting online material This PDF file includes additional material illustrating the express report and supporting its conclusions: Contents: S 1 Satellite image depicting the Waimakariri River catchment and its location within New Zealand S 2 Topographic map of the middle Waimakariri River catchment and its surroundings S 3 Ground images from the study area showing various sites and features mentioned in the text of the express report and supporting the statements presented within (all images: S.Winkler) S 4 Oblique aerial images of the study area (all images: S.Winkler) S 1 Waimakariri River catchment and its location within New Zealand Location of the Waimakariri River catchment within New Zealand. Some other major rivers are in- dicated, also the Waimakariri Gorge (cf. express report, Figure 1; modified after GoogleEarth and (insert) NASA Earth Observatory, https://earthobservatory.nasa.gov). S 2 Middle Waimakariri River catchment and its surroundings Middle Waimakariri River catchment and its surroundings as shown on the NZ250 Topo maps. The area covered by Figure 1 of the express report is indicated by the black frame. -
Discover Arthur's Pass
Discover Arthur's Pass A guide to Arthur's Pass National Park and village Contents Welcome to Arthur's Pass The grandeur of this vast and austere mountain and Welcome to Arthur’s Pass 1 river landscape has instilled awe in those who gaze The historic east-west journey 3 upon it, from the first Māori explorers to modern escapees from city life. Changeable weather—four seasons in one day 8 This once remote area, hidden in the heart of the Dynamic landscape 9 Southern Alps/Kā Tiritiri o Te Moana, was the South Island’s first national park, and is now easily reached Striking plant contrasts from east to west 10 from both Canterbury and the West Coast. Birds at Arthur’s Pass 13 Not only is Arthur’s Pass a key link between east and west, but it is also known for its immense natural Impact of introduced pests and weeds 16 beauty, and rare flora and fauna. The Park provides both a sanctuary for plant and bird life, and a place for Things to do 18 both mental and physical recreation. Since becoming a National Park in 1929, Arthur’s Pass has gained a world- wide reputation for alpine recreation, as well as for its stunning natural history. Published by Department of Conservation Canterbury Conservancy ISBN 978-0-478-14813-8 (Print) Private Bag 4715 ISBN 978-0-478-14814-5 (PDF) Waimakariri River, eastern gateway to the park Photo: G Kates Christchurch 2012 Back cover: Dobson Nature Walk Photo: S Mankelow 1 Arthur’s Pass village Māori inhabitants, to the next wave of immigrants, this time from Europe. -

List of Rivers of New Zealand
Sl. No River Name 1 Aan River 2 Acheron River (Canterbury) 3 Acheron River (Marlborough) 4 Ada River 5 Adams River 6 Ahaura River 7 Ahuriri River 8 Ahuroa River 9 Akatarawa River 10 Akitio River 11 Alexander River 12 Alfred River 13 Allen River 14 Alma River 15 Alph River (Ross Dependency) 16 Anatoki River 17 Anatori River 18 Anaweka River 19 Anne River 20 Anti Crow River 21 Aongatete River 22 Aorangiwai River 23 Aorere River 24 Aparima River 25 Arahura River 26 Arapaoa River 27 Araparera River 28 Arawhata River 29 Arnold River 30 Arnst River 31 Aropaoanui River 32 Arrow River 33 Arthur River 34 Ashburton River / Hakatere 35 Ashley River / Rakahuri 36 Avoca River (Canterbury) 37 Avoca River (Hawke's Bay) 38 Avon River (Canterbury) 39 Avon River (Marlborough) 40 Awakari River 41 Awakino River 42 Awanui River 43 Awarau River 44 Awaroa River 45 Awarua River (Northland) 46 Awarua River (Southland) 47 Awatere River 48 Awatere River (Gisborne) 49 Awhea River 50 Balfour River www.downloadexcelfiles.com 51 Barlow River 52 Barn River 53 Barrier River 54 Baton River 55 Bealey River 56 Beaumont River 57 Beautiful River 58 Bettne River 59 Big Hohonu River 60 Big River (Southland) 61 Big River (Tasman) 62 Big River (West Coast, New Zealand) 63 Big Wainihinihi River 64 Blackwater River 65 Blairich River 66 Blind River 67 Blind River 68 Blue Duck River 69 Blue Grey River 70 Blue River 71 Bluff River 72 Blythe River 73 Bonar River 74 Boulder River 75 Bowen River 76 Boyle River 77 Branch River 78 Broken River 79 Brown Grey River 80 Brown River 81 Buller -

2012 Upper Waimakariri
A Bird survey of the Upper Waimakariri River November 5-8, 2012 J. N. Jolly (On Behalf of BRaid Inc.) May 2013 Abstract A bird survey of 35km of the Upper Waimakariri River (from Bealey Bridge down to the Esk confluence at the top of the gorge) was carried out From November 5-8, 2012 by 14 members of Braided River Aid Inc. (BRaid), with support funding from Environment Canterbury (ECan). With the exception of the black stilt, all the more threatened braided river birds were observed. Previous surveys of the same section of the river were carried out in 1981 and 1995. Wrybill numbers have remained relatively stable, as have banded dotterel, which may even be increasing. Black-fronted terns numbers were lower than in 1995, but much higher than in 1981, while the population of black-billed gulls was much lower than in 1995 but only a little lower than in 1981. The only distinct trends over the three years were Canada geese (upward) and paradise shelduck and duck spp. (downward). Compared to survey results in the Lower Waimakariri, numbers/km of wrybills and banded dotterels were similar, but black- fronted terns and black-billed gulls were lower. In general, bird populations in the Upper Waimakariri were lower than in the upper catchments of the Rangitata and Waitaki (Godley River). However, the results confirm the continuance of the Upper Waimakariri as an important community of riverbed birds both in terms of numbers and diversity. In order to give greater confidence in bird population trends it is recommended that repeat surveys be carried out in 2013 and 2014. -

Draft Canterbury CMS 2013 Vol II: Maps
Map 5.2 Braided Rivers/Ki Uta Ki Tai Place Conservation Management Strategy Canterbury 05 10 20 30 40 Km 15 Reefton Lake Tennyson Waiau River Clarence River Lake Sumner Hurunui South Branch Hurunui River Lake Waiau River Taylor Poulter River Hurunui River Hawdon River Esk River Waimakariri River Ashley River / Rakahuri Waipara River Kowai River Ashley River / Rakahuri Waimakariri River Selwyn River Lake Ellesmere Lake (Te Waihora) Forsyth Akaroa (Wairewa) ASHBURTON Rakaia River Map 5.2.1 Braided Rivers/Ki Uta Ki Tai Place—detail (North) Conservation Management Strategy Canterbury 05 10 20 30 40 Km 16 Lake Hokitika Sumner Hurunui South Branch Hurunui River Lake Taylor Poulter River Hawdon River Bealey River Esk River Waimakariri River Ashley River / Rakahuri Wilberforce River Mathias River Harper River Rakaia River Lake Coleridge Lake Stream Clyde River Lake Havelock River Heron Selwyn River Macaulay River Lake Ellesmere (Te Waihora) Rangitata River ASHBURTON Rakaia River Lake Opuha Ashburton River / Hakatere Hinds River Rangitata River Orari River Opihi River TIMARU Pareora River Map 5.2.2 Braided Rivers/Ki Uta Ki Tai Place—detail (Central) Conservation Management Strategy Canterbury 05 10 20 30 40 Km 17 Mathias River Rakaia River Lake Coleridge Lake Stream Clyde River Lake Havelock River Heron Godley River Macaulay River Murchison River Tasman Lake Rangitata River Tasman River Lake Tekapo Lake Alexandrina Lake Opuha Dobson River Lake Hopkins River Pukaki Tekapo River Lake Orari River Ohau Opihi River Lake Ruataniwha TIMARU Lake Benmore Ahuriri River Pareora River Otaio River Lake Aviemore Lake Waitaki Waimate Waitaki River Map 5.2.3 Braided Rivers/Ki Uta Ki Tai Place—detail (South) Conservation Management Strategy Canterbury 05 10 20 30 40 Km 18.