Kindler Syndrome: Extension of FERMT1 Mutational Spectrum and Natural History
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Revertant Mosaicism in a Human Skin Fragility Disorder Results from Slipped Mispairing and Mitotic Recombination
Revertant mosaicism in a human skin fragility disorder results from slipped mispairing and mitotic recombination Dimitra Kiritsi, … , Leena Bruckner-Tuderman, Cristina Has J Clin Invest. 2012;122(5):1742-1746. https://doi.org/10.1172/JCI61976. Brief Report Dermatology Spontaneous gene repair, also called revertant mosaicism, has been documented in several genetic disorders involving organs that undergo self-regeneration, including the skin. Genetic reversion may occur through different mechanisms, and in a single individual, the mutation can be repaired in various ways. Here we describe a disseminated pattern of revertant mosaicism observed in 6 patients with Kindler syndrome (KS), a genodermatosis caused by loss of kindlin-1 (encoded by FERMT1) and clinically characterized by patchy skin pigmentation and atrophy. All patients presented duplication mutations (c.456dupA and c.676dupC) in FERMT1, and slipped mispairing in direct nucleotide repeats was identified as the reversion mechanism in all investigated revertant skin spots. The sequence around the mutations demonstrated high propensity to mutations, favoring both microinsertions and microdeletions. Additionally, in some revertant patches, mitotic recombination generated areas with homozygous normal keratinocytes. Restoration of kindlin-1 expression led to clinically and structurally normal skin. Since loss of kindlin-1 severely impairs keratinocyte proliferation, we predict that revertant cells have a selective advantage that allows their clonal expansion and, consequently, the improvement of the skin condition. Find the latest version: https://jci.me/61976/pdf Brief report Revertant mosaicism in a human skin fragility disorder results from slipped mispairing and mitotic recombination Dimitra Kiritsi,1 Yinghong He,1 Anna M.G. Pasmooij,2 Meltem Onder,3 Rudolf Happle,1 Marcel F. -

A Review of the Evidence for and Against a Role for Mast Cells in Cutaneous Scarring and Fibrosis
International Journal of Molecular Sciences Review A Review of the Evidence for and against a Role for Mast Cells in Cutaneous Scarring and Fibrosis Traci A. Wilgus 1,*, Sara Ud-Din 2 and Ardeshir Bayat 2,3 1 Department of Pathology, Ohio State University, Columbus, OH 43210, USA 2 Centre for Dermatology Research, NIHR Manchester Biomedical Research Centre, Plastic and Reconstructive Surgery Research, University of Manchester, Manchester M13 9PT, UK; [email protected] (S.U.-D.); [email protected] (A.B.) 3 MRC-SA Wound Healing Unit, Division of Dermatology, University of Cape Town, Observatory, Cape Town 7945, South Africa * Correspondence: [email protected]; Tel.: +1-614-366-8526 Received: 1 October 2020; Accepted: 12 December 2020; Published: 18 December 2020 Abstract: Scars are generated in mature skin as a result of the normal repair process, but the replacement of normal tissue with scar tissue can lead to biomechanical and functional deficiencies in the skin as well as psychological and social issues for patients that negatively affect quality of life. Abnormal scars, such as hypertrophic scars and keloids, and cutaneous fibrosis that develops in diseases such as systemic sclerosis and graft-versus-host disease can be even more challenging for patients. There is a large body of literature suggesting that inflammation promotes the deposition of scar tissue by fibroblasts. Mast cells represent one inflammatory cell type in particular that has been implicated in skin scarring and fibrosis. Most published studies in this area support a pro-fibrotic role for mast cells in the skin, as many mast cell-derived mediators stimulate fibroblast activity and studies generally indicate higher numbers of mast cells and/or mast cell activation in scars and fibrotic skin. -

Foot Pain in Scleroderma
Foot Pain in Scleroderma Dr Begonya Alcacer-Pitarch LMBRU Postdoctoral Research Fellow 20th Anniversary Scleroderma Family Day 16th May 2015 Leeds Institute of Rheumatic and Musculoskeletal Medicine Presentation Content n Introduction n Different types of foot pain n Factors contributing to foot pain n Impact of foot pain on Quality of Life (QoL) Leeds Institute of Rheumatic and Musculoskeletal Medicine Scleroderma n Clinical features of scleroderma – Microvascular (small vessel) and macrovascular (large vessel) damage – Fibrosis of the skin and internal organs – Dysfunction of the immune system n Unknown aetiology n Female to male ratio 4.6 : 1 n The prevalence of SSc in the UK is 8.21 per 100 000 Leeds Institute of Rheumatic and Musculoskeletal Medicine Foot Involvement in SSc n Clinically 90% of SSc patients have foot involvement n It typically has a later involvement than hands n Foot involvement is less frequent than hand involvement, but is potentially disabling Leeds Institute of Rheumatic and Musculoskeletal Medicine Different Types of Foot Pain Leeds Institute of Rheumatic and Musculoskeletal Medicine Ischaemic Pain (vascular) Microvascular disease (small vessel) n Intermittent pain – Raynaud’s (spasm) • Cold • Throb • Numb • Tingle • Pain n Constant pain – Vessel center narrows • Distal pain (toes) • Gradually increasing pain • Intolerable pain when necrosis is present Leeds Institute of Rheumatic and Musculoskeletal Medicine Ischaemic Pain (vascular) Macrovascular disease (large vessels) n Intermittent and constant pain – Peripheral Arterial Disease • Intermittent claudication – Muscle pain (ache, cramp) during walking • Aching or burning pain • Night and rest pain • Cramps Leeds Institute of Rheumatic and Musculoskeletal Medicine Ulcer Pain n Ulcer development – Constant pain n Infected ulcer – Unexpected/ excess pain or tenderness Leeds Institute of Rheumatic and Musculoskeletal Medicine Neuropathic Pain n Nerve damage is not always obvious. -
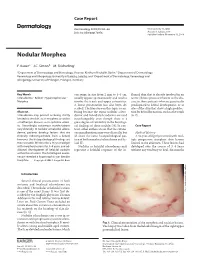
Nodular Morphea
Case Report Dermatology 2009;218:63–66 Received: July 13, 2008 DOI: 10.1159/000173976 Accepted: July 23, 2008 Published online: November 13, 2008 Nodular Morphea a b c F. Kauer J.C. Simon M. Sticherling a b Department of Dermatology and Venerology, Vivantes Klinikum Neukölln, Berlin , Department of Dermatology, c Venerology and Allergology, University of Leipzig, Leipzig , and Department of Dermatology, Venerology and Allergology, University of Erlangen, Erlangen , Germany Key Words can range in size from 2 mm to 4–5 cm, flamed skin that is already involved in an -Scleroderma ؒ Keloid ؒ Hypertrophic scar ؒ usually appear spontaneously and tend to active fibrotic process inherent to the dis Morphea involve the trunk and upper extremities. ease in those patients who are genetically A linear presentation has also been de- predisposed to keloid development, or at scribed. The literature on this topic is con- sites of the skin that show a high predilec- Abstract fusing because the terms ‘nodular sclero- tion for keloid formation, such as the trunk Scleroderma may present as being strictly derma’ and ‘keloidal scleroderma’ are used [6, 7] . limited to the skin, as in morphea, or within interchangeably even though there is a a multiorgan disease, as in systemic sclero- great degree of variability in the histologi- sis. Accordingly, cutaneous manifestations cal findings of these nodules [4] . In con- C a s e R e p o r t vary clinically. In nodular or keloidal sclero- trast, other authors stress that the cutane- derma, patients develop lesions that are ous manifestations may vary clinically, but Medical History clinically indistinguishable from a keloid; all share the same histopathological pat- A 16-year-old girl presented with mul- however, the histopathological findings are tern of both morphea/scleroderma and ke- tiple progressive morpheic skin lesions more variable. -

Topical Photodynamic Therapy for Localized Scleroderma
Acta Derm Venereol 2000; 80: 26±27 Topical Photodynamic Therapy for Localized Scleroderma SIGRID KARRER, CHRISTOPH ABELS, MICHAEL LANDTHALER and ROLF-MARKUS SZEIMIES Department of Dermatology, University of Regensburg, Regensburg, Germany Therapy of localized scleroderma is unsatisfactory, with MATERIAL AND METHODS numerous treatments being used that have only limited success Patients or considerable side-effects. The aim of this trial was to determine whether topical photodynamic therapy would be Five patients aged between 21 and 63 years with biopsy-proven effective in patients with localized scleroderma. Five patients localized scleroderma (morphoea) were studied. All patients had evidence of progressive disease, i.e. lesions showed an in¯ammatory with progressive disease, in whom conventional therapies had reaction and increase in size. In each patient the condition had failed, were treated by application of a gel containing 3% previously failed to respond to potent topical corticosteroids, systemic 5-aminolevulinic acid followed by irradiation with an incoherent therapy with penicillin and/or PUVA bath photochemotherapy. lamp (40 mW/cm2, 10 J/cm2). The treatment was performed Patients were required to discontinue all therapy at least 4 weeks once or twice weekly for 3 ± 6 months. In all patients the before study initiation. The most affected sclerotic plaque of each therapy was highly effective for sclerotic plaques, as measured patient was judged at baseline and then every 2 weeks during by a quantitative durometer score and a clinical skin score. The treatment. Sclerotic plaques were assessed by a clinical skin score (10) only side-effect was a transient hyperpigmentation of the and the durometer score (11). -

Dumitras, Cu, MC; Popa, A.; Petca, A.; Miulescu, R.-G. Cutaneous Mastocytosis in Childhood—Update from the Literature
Journal of Clinical Medicine Review Cutaneous Mastocytosis in Childhood—Update from the Literature 1,2 1,3 1,4 1,5, Florica Sandru ,Răzvan-Cosmin Petca , Monica Costescu , Mihai Cristian Dumitras, cu *, Adelina Popa 2, Aida Petca 1,6,* and Raluca-Gabriela Miulescu 1,7 1 “Carol Davila” University of Medicine and Pharmacy, 030167 Bucharest, Romania; fl[email protected] (F.S.); [email protected] (R.-C.P.); [email protected] (M.C.); [email protected] (R.-G.M.) 2 Department of Dermatology, Elias University Emergency Hospital, 0611461 Bucharest, Romania; [email protected] 3 Department of Urology, “Prof. Dr. Theodor Burghele” Clinical Hospital, 061344 Bucharest, Romania 4 Department of Dermatology, “Dr. Victor Babes” Clinical Hospital of Infectious and Tropical Diseases, 030303 Bucharest, Romania 5 Department of Obstetrics & Gynecology, University Emergency Hospital, 050098 Bucharest, Romania 6 Department of Obstetrics & Gynecology, Elias University Emergency Hospital, 0611461 Bucharest, Romania 7 Department of Dermatology, Vălenii de Munte Hospital, 106400 Prahova, Romania * Correspondence: [email protected] (M.C.D.); [email protected] (A.P.); Tel.: +40-722-223223 (M.C.D.); +40-745-787448 (A.P.) Abstract: Mastocytosis (M) represents a systemic pathology characterized by increased accumulation and clonal proliferation of mast cells in the skin and/or different organs. Broadly, M is classified into two categories: Cutaneous mastocytosis (CM) and systemic mastocytosis (SM). In children, CM is Citation: Sandru, F.; Petca, R.-C.; the most frequent form. Unfortunately, pathogenesis is still unclear. It is thought that genetic factors Costescu, M.; Dumitras, cu, M.C.; are involved, but further studies are necessary. As for features of CM, the lesions differ in clinical Popa, A.; Petca, A.; Miulescu, R.-G. -

Genodermatoses
GENODERMATOSES Genodermatoses What’s new? Nigel P Burrows C Filaggrin mutations underlie ichthyosis vulgaris and are a risk factor for atopy including eczema, allergic sensitization, asthma, allergic rhinitis and peanut allergy Abstract C A new classification and nomenclature for ichthyoses was Genetic skin diseases encompass a spectrum from the common to the published in 2009, peeling skin syndromes which may be rare. It is important for the clinician to be alert to the possibility that confused for the milder subtypes of epidermolysis bullosa are the patient may be presenting for the first time with one or more features distinct genetic entities of a genetic disease so that appropriate investigation and counselling can C Emerging evidence that pseudoxanthoma elasticum is a meta- take place. Recent discoveries have helped the understanding of many of bolic disorder resulting in calcification of elastic fibres these disorders. A few common and important genodermatoses are high- C Mammalian target of rapamycin (mTOR) inhibitor therapies are lighted in this article. showing promise in tuberous sclerosis complex C Vascular anomalies on the skin may be the presenting feature of Keywords cancer syndromes; collagen; epidermolysis bullosa; filaggrin; inherited syndromes genodermatoses; ichthyosis; keratinization; pseudoxanthoma elasticum; vascular anomalies X-linked recessive ichthyosis In 75% of cases of X-linked recessive ichthyosis (XLRI) (Figure 1), scaling is present in the first week of life and tends to progress into adolescence. In contrast to IV, the flexures may be Genetic skin diseases encompass a spectrum from the common involved. A third of cases are associated with a prolonged labour. (e.g. atopic eczema) to the rare (e.g. -

Oral Health Care for Patients with Epidermolysis Bullosa
Oral Health Care for Patients with Epidermolysis Bullosa Best Clinical Practice Guidelines October 2011 Oral Health Care for Patients with Epidermolysis Bullosa Best Clinical Practice Guidelines October 2011 Clinical Editor: Susanne Krämer S. Methodological Editor: Julio Villanueva M. Authors: Prof. Dr. Susanne Krämer Dr. María Concepción Serrano Prof. Dr. Gisela Zillmann Dr. Pablo Gálvez Prof. Dr. Julio Villanueva Dr. Ignacio Araya Dr. Romina Brignardello-Petersen Dr. Alonso Carrasco-Labra Prof. Dr. Marco Cornejo Mr. Patricio Oliva Dr. Nicolás Yanine Patient representatives: Mr. John Dart Mr. Scott O’Sullivan Pilot: Dr. Victoria Clark Dr. Gabriela Scagnet Dr. Mariana Armada Dr. Adela Stepanska Dr. Renata Gaillyova Dr. Sylvia Stepanska Review: Prof. Dr. Tim Wright Dr. Marie Callen Dr. Carol Mason Prof. Dr. Stephen Porter Dr. Nina Skogedal Dr. Kari Storhaug Dr. Reinhard Schilke Dr. Anne W Lucky Ms. Lesley Haynes Ms. Lynne Hubbard Mr. Christian Fingerhuth Graphic design: Ms. Isabel López Production: Gráfica Metropolitana Funding: DEBRA UK © DEBRA International This work is subject to copyright. ISBN-978-956-9108-00-6 Versión On line: ISBN 978-956-9108-01-3 Printed in Chile in October 2011 Editorial: DEBRA Chile Acknowledgement: We would like to thank Coni V., María Elena, María José, Daniela, Annays, Lisette, Victor, Coni S., Esteban, Coni A., Felipe, Nibaldo, María, Cristián, Deyanira and Victoria for sharing their smile to make these Guidelines more friendly. 4 Contents 1 Introduction 07 2 Oral care for patients with Inherited Epidermolysis Bullosa 11 3 Dental treatment 19 4 Anaesthetic management 29 5 Summary of recommendations 33 Development of the guideline 37 6 Appendix 43 7.1 List of abbreviations and glossary 7.2 Oral manifestations of Epidermolysis Bullosa 7 7.3 General information on Epidermolysis Bullosa 7.4 Exercises for mouth, jaw and tongue 8 References 61 5 A message from the patient representative: “Be guided by the professionals. -

Immune Thrombocytopenic Purpura (ITP) — Adult Conditions for Which Ivig Has an Established Therapeutic Role
Immune thrombocytopenic purpura (ITP) — adult Conditions for which IVIg has an established therapeutic role. Specific Conditions Newly Diagnosed Immune thrombocytopenic purpura (ITP) Persistent Immune thrombocytopenic purpura (ITP) Chronic Immune thrombocytopenic purpura (ITP) Evans syndrome ‐ with significant Immune thrombocytopenic purpura (ITP) ‐ adult Indication for IVIg Use Newly diagnosed ITP — initial Ig therapy ITP in pregnancy — initial Ig therapy ITP with life‐threatening haemorrhage or the potential for life‐threatening haemorrhage Newly diagnosed or persistent ITP — subsequent therapy (diagnosis <12 months) Refractory persistent or chronic ITP — splenectomy failed or contraindicated and second‐line agent unsuccessful Subsequent or ongoing treatment for ITP responders during pregnancy and the postpartum period ITP and inadequate platelet count for planned surgery HIV‐associated ITP Level of Evidence Evidence of probable benefit – more research needed (Category 2a) Description and Diagnostic Immune thrombocytopenic purpura (ITP) is a reduction in platelet count Criteria (thrombocytopenia) resulting from shortened platelet survival due to anti‐platelet antibodies, reduced platelet production due to immune induced reduced megakaryopoeisis and/or immune mediated direct platelet lysis. When counts are very low (less than 30x109/L), bleeding into the skin (purpura) and mucous membranes can occur. Bone marrow platelet production (megakaryopoiesis) is morphologically normal. In some cases, there is additional impairment of platelet function related to antibody binding to glycoproteins on the platelet surface. It is a common finding in patients with human immunodeficiency virus (HIV) disease, and while it may be found at any stage of the infection, its prevalence increases as HIV disease advances. Around 80 percent of adults with ITP have the chronic form of disease. -

Systemic Sclerosis/Scleroderma: a Treatable Multisystem Disease
Systemic Sclerosis/Scleroderma: A Treatable Multisystem Disease MONIQUE HINCHCLIFF, MD, and JOHN VARGA, MD Northwestern University, Feinberg School of Medicine, Chicago, Illinois Systemic sclerosis (systemic scleroderma) is a chronic connective tissue disease of unknown etiology that causes wide- spread microvascular damage and excessive deposition of collagen in the skin and internal organs. Raynaud phenome- non and scleroderma (hardening of the skin) are hallmarks of the disease. The typical patient is a young or middle-age woman with a history of Raynaud phenomenon who presents with skin induration and internal organ dysfunction. Clinical evaluation and laboratory testing, along with pulmonary function testing, Doppler echocardiography, and high-resolution computed tomography of the chest, establish the diagnosis and detect visceral involvement. Patients with systemic sclerosis can be classified into two distinct clinical subsets with different patterns of skin and internal organ involvement, autoantibody production, and survival. Prognosis is determined by the degree of internal organ involvement. Although no disease-modifying therapy has been proven effective, complications of systemic sclerosis are treatable, and interventions for organ-specific manifestations have improved substantially. Medications (e.g., cal- cium channel blockers and angiotensin-II receptor blockers for Raynaud phenomenon, appropriate treatments for gastroesophageal reflux disease) and lifestyle modifications can help prevent complications, such as digital ulcers and Barrett esophagus. Endothelin-1 receptor blockers and phosphodiesterase-5 inhibitors improve pulmonary arte- rial hypertension. The risk of renal damage from scleroderma renal crisis can be lessened by early detection, prompt initiation of angiotensin-converting enzyme inhibitor therapy, and avoidance of high-dose corticosteroids. Optimal patient care includes an integrated, multidisciplinary approach to promptly and effectively recognize, evaluate, and manage complications and limit end-organ dysfunction. -

Genevista Epidermolysis Bullosa
GeNeViSTA Epidermolysis Bullosa: An Update Neetu Bhari1, Neerja Gupta2 1Department of Dermatology and Venereology, All India Institute of Medical Sciences, New Delhi, India 2Division of Genetics, Department of Pediatrics, All India Institute of Medical Sciences, New Delhi, India Correspondence to: Dr Neerja Gupta Email: [email protected] Abstract Classification and pathogenesis Epidermolysis bullosa (EB) is caused by mutation in Mutation in various components of the basement various components of the basement membrane membrane zone responsible for structural support zone and is characterized by increased fragility of in keratinocytes and adhesion of epidermis and skin and mucous membrane. There are four major dermis, results in different types of epidermol- types based on the level of split at the basement ysis bullosa (Figure 1). The new approach for membrane zone – involvement of basal layer of classification in EB is termed as “onion skinning” epidermis in epidermolysis bullosa simplex (EBS), and sequentially takes account of the level of split lamina lucida in junctional EB (JEB) and sublamina at the dermato-epidermal basement membrane densa in dystrophic EB (DEB) while Kindler syn- zone, phenotypic features and genetic characteris- drome (KS) can exhibit multiple levels of split. More tics such as mode of inheritance, targeted protein than 19 genes have been described resulting in the and its relative expression in the basement mem- various subtypes of these 4 major types giving rise brane zone, the involved gene and the type of to diversity of phenotypic expressions. Diagnosis mutation (Fine et al., 2014). is based on the “onion skin approach” which in- The level of split is the basal layer of the cludes clinical presentation, family history, antigen epidermis in epidermolysis bullosa simplex (EBS) mapping and genetic mutation analysis. -

How to Deal with Skin Biopsy in an Infant with Blisters?
Review How to Deal with Skin Biopsy in an Infant with Blisters? Stéphanie Leclerc-Mercier Reference Center for Genodermatoses (MAGEC Center), Department of Pathology, Necker-Enfants Malades Hospital, Paris Centre University, 75015 Paris, France; [email protected] Abstract: The onset of blisters in a neonate or an infant is often a source of great concern for both parents and physicians. A blistering rash can reveal a wide range of diseases with various backgrounds (infectious, genetic, autoimmune, drug-related, traumatic, etc.), so the challenge for the dermatologist and the pediatrician is to quickly determine the etiology, between benign causes and life-threatening disorders, for a better management of the patient. Clinical presentation can provide orientation for the diagnosis, but skin biopsy is often necessary in determining the cause of blister formations. In this article, we will provide information on the skin biopsy technique and discuss the clinical orientation in the case of a neonate or infant with a blistering eruption, with a focus on the histology for each etiology. Keywords: blistering eruption; infant; skin biopsy; genodermatosis; SSSS; hereditary epidermolysis bul- losa; keratinopathic ichthyosis; incontinentia pigmenti; mastocytosis; auto-immune blistering diseases 1. Introduction The onset of blisters in a neonate or an infant (<2 years old) is a source of great concern for both parents and physicians. Therefore, a precise diagnosis, between benign causes and Citation: Leclerc-Mercier, S. How to life-threatening disorders, is quickly needed for the best management of the baby. Deal with Skin Biopsy in an Infant Several diseases with various backgrounds (infectious, genetic, autoimmune, drug- with Blisters? Dermatopathology 2021, related, traumatic, etc.) can lead to a blistering eruption.