A PRELIMINARY STUDY of FRUIT HANDLING by CAPTIVE BORNEAN ORANGUTANS Pongo Pygmaeus Pygmaeus and the EFFECTS on SEED GERMINATION
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Five Hundred Plant Species in Gunung Halimun Salak National Park, West Java a Checklist Including Sundanese Names, Distribution and Use
Five hundred plant species in Gunung Halimun Salak National Park, West Java A checklist including Sundanese names, distribution and use Hari Priyadi Gen Takao Irma Rahmawati Bambang Supriyanto Wim Ikbal Nursal Ismail Rahman Five hundred plant species in Gunung Halimun Salak National Park, West Java A checklist including Sundanese names, distribution and use Hari Priyadi Gen Takao Irma Rahmawati Bambang Supriyanto Wim Ikbal Nursal Ismail Rahman © 2010 Center for International Forestry Research. All rights reserved. Printed in Indonesia ISBN: 978-602-8693-22-6 Priyadi, H., Takao, G., Rahmawati, I., Supriyanto, B., Ikbal Nursal, W. and Rahman, I. 2010 Five hundred plant species in Gunung Halimun Salak National Park, West Java: a checklist including Sundanese names, distribution and use. CIFOR, Bogor, Indonesia. Photo credit: Hari Priyadi Layout: Rahadian Danil CIFOR Jl. CIFOR, Situ Gede Bogor Barat 16115 Indonesia T +62 (251) 8622-622 F +62 (251) 8622-100 E [email protected] www.cifor.cgiar.org Center for International Forestry Research (CIFOR) CIFOR advances human wellbeing, environmental conservation and equity by conducting research to inform policies and practices that affect forests in developing countries. CIFOR is one of 15 centres within the Consultative Group on International Agricultural Research (CGIAR). CIFOR’s headquarters are in Bogor, Indonesia. It also has offices in Asia, Africa and South America. | iii Contents Author biographies iv Background v How to use this guide vii Species checklist 1 Index of Sundanese names 159 Index of Latin names 166 References 179 iv | Author biographies Hari Priyadi is a research officer at CIFOR and a doctoral candidate funded by the Fonaso Erasmus Mundus programme of the European Union at Southern Swedish Forest Research Centre, Swedish University of Agricultural Sciences. -

Collection and Evaluation of Under-Utilized Tropical and Subtropical Fruit Tree Genetic Resources in Malaysia
J]RCAS International Symposium Series No. 3: 27-38 Session 1-3 27 Collection and Evaluation of Under-Utilized Tropical and Subtropical Fruit Tree Genetic Resources in Malaysia WONG, Kai Choo' Abstract Fruit tree genetic resources in Malaysia consist of cultivated and wild species. The cul tivated fruit trees number more than 100 species of both indigenous and introduced species. Among these fruits, some are popular and are widely cultivated throughout the country while others are less known and grown in small localized areas. The latter are the under-utilized fruit species. Apart from these cultivated fruits, there is also in the Malaysian natural forest a diversity of wild fruit tree species which produce edible fruits but are relatively unknown and unutilized. Many of the under-utilized and unutilized fruit species are known to show economic potential. Collection and evaluation of some of these fruit tree genetic resources have been carried out. These materials are assessed for their potential as new fruit trees, as sources of rootstocks for grafting and also as sources of germplasm for breeding to improve the present cultivated fruit species. Some of these potential fruit tree species within the gen era Artocarpus, Baccaurea, Canarium, Dimocarpus, Dialium, Durio, Garcinia, Litsea, Mangif era, Nephelium, Sa/acca, and Syzygium are highlighted. Introduction Malaysian fruit tree genetic resources comprise both cultivated and wild species. There are more than 100 cultivated fruit species of both major and minor fruit crops. Each category includes indigenous as well as introduced species. The major cultivated fruit crops are well known and are commonly grown throughout the country. -

Ecological Assessments in the B+WISER Sites
Ecological Assessments in the B+WISER Sites (Northern Sierra Madre Natural Park, Upper Marikina-Kaliwa Forest Reserve, Bago River Watershed and Forest Reserve, Naujan Lake National Park and Subwatersheds, Mt. Kitanglad Range Natural Park and Mt. Apo Natural Park) Philippines Biodiversity & Watersheds Improved for Stronger Economy & Ecosystem Resilience (B+WISER) 23 March 2015 This publication was produced for review by the United States Agency for International Development. It was prepared by Chemonics International Inc. The Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience Program is funded by the USAID, Contract No. AID-492-C-13-00002 and implemented by Chemonics International in association with: Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) The author’s views expressed in this publication do not necessarily reflect the views of the United States Agency for International Development or the United States Government. Ecological Assessments in the B+WISER Sites Philippines Biodiversity and Watersheds Improved for Stronger Economy and Ecosystem Resilience (B+WISER) Program Implemented with: Department of Environment and Natural Resources Other National Government Agencies Local Government Units and Agencies Supported by: United States Agency for International Development Contract No.: AID-492-C-13-00002 Managed by: Chemonics International Inc. in partnership with Fauna and Flora International (FFI) Haribon Foundation World Agroforestry Center (ICRAF) 23 March -

Local and Landscape Compositions Influence Stingless Bee Communities and Pollination Networks in Tropical Mixed Fruit Orchards
diversity Article Local and Landscape Compositions Influence Stingless Bee Communities and Pollination Networks in Tropical Mixed Fruit Orchards, Thailand Kanuengnit Wayo 1 , Tuanjit Sritongchuay 2 , Bajaree Chuttong 3, Korrawat Attasopa 3 and Sara Bumrungsri 1,* 1 Department of Biology, Division of Biological Science, Faculty of Science, Prince of Songkla University, Hat Yai, Songkhla 90110, Thailand; [email protected] 2 Center for Integrative Conservation, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Menglun, Mengla 666303, China; [email protected] 3 Department of Entomology and Plant Pathology, Faculty of Agriculture, Chiang Mai University, Chiang Mai 50200, Thailand; [email protected] (B.C.); [email protected] (K.A.) * Correspondence: [email protected]; Tel.: +66-7428-8537 Received: 31 October 2020; Accepted: 15 December 2020; Published: 17 December 2020 Abstract: Stingless bees are vital pollinators for both wild and crop plants, yet their communities have been affected and altered by anthropogenic land-use change. Additionally, few studies have directly addressed the consequences of land-use change for meliponines, and knowledge on how their communities change across gradients in surrounding landscape cover remains scarce. Here, we examine both how local and landscape-level compositions as well as forest proximity affect both meliponine species richness and abundance together with pollination networks across 30 mixed fruit orchards in Southern Thailand. The results reveal that most landscape-level factors significantly influenced both stingless bee richness and abundance. Surrounding forest cover has a strong positive direct effect on both factors, while agricultural and urbanized cover generally reduced both bee abundance and diversity. In the local habitat, there is a significant interaction between orchard size and floral richness with stingless bee richness. -
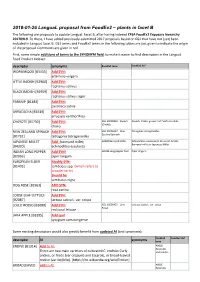
2018-01-26 Langual Proposal from Foodex2 – Plants in Facet B
2018-01-26 LanguaL proposal from FoodEx2 – plants in facet B The following are proposals to update LanguaL Facet B, after having indexed EFSA FoodEx2 Exposure hierarchy 20170919. To these, I have added previously-submitted 2017 proposals based on GS1 that have not (yet) been included in LanguaL facet B. GS1 terms and FoodEx2 terms in the following tables are just given to indicate the origin of the proposal. Comments are given in red. First, some simple additions of terms to the SYNONYM field, to make it easier to find descriptors in the LanguaL Food Product Indexer: descriptor synonyms FoodEx2 term FoodEx2 def WORMWOOD [B3433] Add SYN: artemisia vulgaris LITTLE RADISH [B2960] Add SYN: raphanus sativus BLACK RADISH [B2959] Add SYN: raphanus sativus niger PARSNIP [B1483] Add SYN: pastinaca sativa ARRACACHA [B3439] Add SYN: arracacia xanthorrhiza CHAYOTE [B1730] Add SYN: GS1 10006356 - Squash Squash, Choko, grown from Sechium edule (Choko) choko NEW ZEALAND SPINACH Add SYN: GS1 10006427 - New- Tetragonia tetragonoides Zealand Spinach [B1732] tetragonia tetragonoides JAPANESE MILLET Add : barnyard millet; A000Z Barnyard millet Echinochloa esculenta (A. Braun) H. Scholz, Barnyard millet or Japanese Millet. [B4320] echinochloa esculenta INDIAN LONG PEPPER Add SYN! A019B Long pepper fruit Piper longum [B2956] piper longum EUROPEAN ELDER Modify SYN: [B1403] sambucus spp. (which refers to broader term) Should be sambucus nigra DOG ROSE [B2961] ADD SYN: rosa canina LOOSE LEAF LETTUCE Add SYN: [B2087] lactusa sativa L. var. crispa LOLLO ROSSO [B2088] Add SYN: GS1 10006425 - Lollo Lactuca sativa L. var. crispa Rosso red coral lettuce JAVA APPLE [B3395] Add syn! syzygium samarangense Some existing descriptors would also greatly benefit from updated AI (and synonyms): FoodEx2 FoodEx2 def descriptor AI synonyms term ENDIVE [B1314] Add to AI: A00LD Escaroles There are two main varieties of cultivated C. -

Condition of Approved Fruits Rambutan, Durian, Mangosteen, Longan, Lychee, Pomegranate, and Jackfruit for Certification for Movement Interstate
INTERSTATE CERTIFICATION ASSURANCE ICA-13 CONDITION OF APPROVED FRUITS RAMBUTAN, DURIAN, MANGOSTEEN, LONGAN, LYCHEE, POMEGRANATE, AND JACKFRUIT FOR CERTIFICATION FOR MOVEMENT INTERSTATE REVISION REGISTER Revision No. Date of Issue Amendment Details Third 26/10/1998 All Pages Fourth 18/12/2002 Pages 3, 4, 5, 6, 7, 8, 10, 12, 15, 16 Fifth 27/01/2005 Pages 13 & 14 Sixth 26/07/2005 All Pages Seventh 03/06/2010 All Pages Eighth 21/05/2014 All Pages Ninth 21/06/2016 Updated Issue Controlled Copy No: _____________________ Controlled: Authorised: _____________________ Uncontrolled: Quality Control Officer Uncontrolled copies of this document are not subject to amendment or revision. Holders of uncontrolled copies should contact the DPIR, Operations Manager, PBB, GPO Box 3000, Darwin NT 0801, Telephone: (08) 8999 2118 to obtain a current version of this document. OPERATIONAL PROCEDURE - ICA13 PAGE 1 OF 23 PAGES TABLE OF CONTENTS 1. PURPOSE ........................................................................................................................... 4 2. SCOPE ................................................................................................................................ 4 3. REFERENCES .................................................................................................................... 4 4. DEFINITIONS ...................................................................................................................... 4 5. RESPONSIBILITY .............................................................................................................. -

Foods to Share Family Style
Soups Beef & Pork (32 oz) Beef Broccoli ....................................................................9.95 Egg Drop Soup w. Sweet Corn & Minced Chicken...............7.95 Beef w. Choy Sum ..........................................................10.95 Sinigang w. Shrimp ..........................................................9.95 Wok Stir-fry Vegetables w. Beef .....................................10.95 Bitter Melon w. Beef ......................................................11.95 Appetizers Foods to Share Mongolian Beef .............................................................11.95 Crispy Gau Gee............................................................each .60 Sweet & Sour Pork ...........................................................9.95 Vegetable Lumpia ....................................................each 1.00 Family Style Kung Pao Beef ...............................................................11.95 Shanghai Lumpia .....................................................each 1.00 Shrimp & Fish Teriyaki Beef 12 oz ..........................................................13.95 Gyoza (4 pc.) .....................................................................4.25 Stir-fry Garlic Shrimp Kau Yuk (Pot Roast Pork) .................................................10.95 Buffalo Chicken Wing (5 pc.) .............................................6.95 w. Vegetables ............................................................12.95 Sweet & Sour Ribs .........................................................10.95 Crispy Seasoned -

Study on Measurement and Determination Carbon Pool in Traditional Agroforestry System for Handling Climate Change
International Journal of Forestry and Horticulture (IJFH) Volume 4, Issue 2, 2018, PP 14-24 ISSN No. (Online) 2454–9487 DOI: http://dx.doi.org/10.20431/2454-9487.0402003 www.arcjournals.org Study on Measurement and Determination Carbon Pool in Traditional Agroforestry System for Handling Climate Change Jan Willem Hatulesila1, Cornelia M.A.Wattimena1, Ludia Siahaya1 1Forestry Department, Faculty of Agriculture, Pattimura University *Corresponding Author: Jan Willem Hatulesila, Forestry Department, Faculty of Agriculture, Pattimura University. Abstract: The forest that is the lungs of the world plays a very important role in the continuity of life on earth. Forests absorb Co2 during the process of photosynthesis and decomposition as organic matter in plant biomass. Measurement of forest productivity by biomass measurement. Forest biomass provides important information in the predicted potential of Co2 and biomass sequestration in certain ages that can be used to estimate forest productivity. This research was conducted in the community forest of Hutumuri village as one of traditional agroforestry “dusung” in Ambon Island. Beside that data base can serve as a planner base for the development of large-scale forecasts of carbon stocks for total community forest of agroforestry patterns. The carbon measurement method is done in demonstration plot for tree, sapling and forest floor level. The results show the dominant plant species for plot I in the dominance of nutmeg (Myristica fragrans), plot II durian (Durio zibethinus) and plot III duku (Lansium spp). The total measured biomass on tree vegetation was 58.52 tC / ha; forest floor was 1.71 tC / ha; top soil layer was 13 tC / ha, woody necromass was 33.56 kg 2 / m ; rough and soft litter were 1.84 tC / ha, necromass was 67.16 tC / ha. -

Thai Export of Durian to China
Impact of China’s Increasing Demand for Agro Produce on Agricultural Production in the Mekong Region, BRC Research Report Bangkok Research Center, JETRO Bangkok/IDE-JETRO, 2018 Chapter 5 Thai Export of Durian to China Wannarat Tantrakonnsab1 and Nattapon Tantrakoonsab2 Abstract In the last five years or longer, exports of Thai durian has been increasing consistently. Although other fruit exported from Thailand also travels in the same direction, durian is the outstanding fruit dominating the China market. There are several factors that affect this development, including the Chinese regulations, development of the durian industry in Thailand, the role of Chinese entrepreneurs, and transport improvements in the Mekong region's countries. Meanwhile the increase of Thai durian exports to China has created changes for Thai durian exports. The durian industry has expanded through high consumption, and some exporters' business activities have enlarged, some have disappeared, or been replaced by Chinese entrepreneurs. Although some Thai stakeholders in the process, such as the farmers and sorting and packing plants, may benefit from the higher export volume of durian at present, there are still some concerns that they may face in the future. 1. Durian, and the increasing export of Thai fruit to China Before the political reforms and open trading policy of China since the end of the 1970s or around thirty or forty years ago, Thai fruit was shipped to Hong Kong as the main 1 Institute of Asian Studies, Chulalongkorn University, Thailand 2 Institute of Asian Studies, Chulalongkorn University, Thailand 1 Impact of China’s Increasing Demand for Agro Produce on Agricultural Production in the Mekong Region, BRC Research Report Bangkok Research Center, JETRO Bangkok/IDE-JETRO, 2018 market, and only some was re-exported to China. -

Chapter 3 Figure 7
Cover Page The handle http://hdl.handle.net/1887/138409 holds various files of this Leiden University dissertation. Author: Darmanto Title: Good to produce, good to share: Food, hunger, and social values in a contemporary Mentawaian community, Indonesia Issue date: 2020-11-26 3 Plenty of Kat, Lack of Iba: The Availability of and Access to Edible Resources This chapter will describe and present a variety of ecological and quantitative data to demonstrate the availability and the access to food resources around the settlement. This chapter starts with the emic category of ecosystem. Understanding how people use the natural environment and create specific zones will enable us to examine the complex relations between Muntei residents and the environment, the pattern of resources’ exploitation, and in particular, food availability and the access to food resources. The following sections will describe the availability of and the access to a wide range of food resources from plants and animals as well as imported food. 3.1 Manipulated Ecosystem and Productive Zones People see the environment around their settlement as a vast resource containing edible and non-edible animals and plants, which can be exploited. They have established a tenure arrangement in which empty territory and unclaimed objects on it did not exist. All the land, rivers, caves, waterfalls, small lakes, mangroves, and other specific ecosystems have been occupied, named, claimed, and exploited. Each ecosystem and the resources on it are far from uniform, for reasons of their physical and natural character. The variations within those ecosystems are quite considerable, determining the ways people make use of them and the breadth of choice available in the selection of edible resources. -

Lepidoptera, Cossidae) from Indonesia and Thailand
Ecologica Montenegrina 38: 102-106 (2020) This journal is available online at: www.biotaxa.org/em http://dx.doi.org/10.37828/em.2020.38.12 https://zoobank.org/urn:lsid:zoobank.org:pub:05A73DF2-AC30-4CDD-A01E-E1999D5B1A3D Two new species of Roepkiella Yakovlev & Witt, 2009 (Lepidoptera, Cossidae) from Indonesia and Thailand ROMAN V. YAKOVLEV Altai State University, pr. Lenina 61, Barnaul, 656049, Russia. E-mail: [email protected] Tomsk State University, Laboratory of Biodiversity and Ecology, Lenina pr. 36, 634050 Tomsk, Russia Received 21 November 2020 │ Accepted by V. Pešić: 9 December 2020 │ Published online 10 December 2020. Abstract The article describes two new species: Roepkiella jakli sp. nov. (type locality: Indonesia, Sangir Isl., Tahuna Distr., Bukit Bembalut Hill) and R. korshunovi sp. nov. (type locality: Thailand, Khon Kaen Prov., Phu Wiang Wat). The diagnostic features are given, the male genitalia of R. celebensis (Roepke, 1957) are described for the first time. Key words: biodiversity, Cossoidea, entomology, Asia, Paleotropics, Oriental Region, taxonomy, new species. Introduction The genus Roepkiella Yakovlev & Witt, 2009 (Lepidoptera, Cossidae) was firstly described for Cossus rufidorsia Hampson, 1904 (type locality: Sikhim [E. India, Sikkim prov.]) by original designation (Yakovlev & Witt 2009). According to the publications (Roepke 1957; Arora 1976; Holloway 1986; Schoorl 1990; Yakovlev 2011, 2014) the genus includes 14 species, distributed in Hindustan, Indochina, Myanmar, Malacca, Borneo, Java, Sumatra, and Sulawesi. Their biology is very poorly studied. Only for R. chlorata (Swinhoe, 1892) (type locality: Sarawak, Borneo) and Roepkiella subfusca (Snellen, 1895) (type locality: [Deli, Ost Sumatra]) the feed plants were indicated: Intsia palembanica Miquel, 1861, Parkia sp. -

Report- Phase I
ENVIRONMENTAL IMPACT ASSESSMENT REPORT- PHASE I September 2020 DRAFT EIA REPORT Bangalore – Chennai Expressway-Phase-I TOR Compliances The following are the compliances for the ToR issued by MOEF vide letter NO.F.No. 10-15/2018- IA.III dated 14th May 2018 for the Bangalore-Chennai Expressway Phase-I. Sr. No. TOR Points & Conditions Compliances A Project Specific Conditions Quantity and Source of water utilization and its Section 4.2.3.3 of Chapter-4 i permission ii Traffic density study to be carried out. Section 2.7.3 of Chapter-2 Cumulative impact assessment study to be carried Refer Section 4.5 of Chapter 4 iii out along proposed alignment including other packages. Detailed Impact assessment study to be carried out Chapter-4 and Chapter-9 iv with mitigation measures for protection of reservoirs, water bodies and rivers if any Composite plan to compensate the loss of tree cover Annexure-9.1 of Chapter-9 v though massive tree plantation programme with time schedule and financial outlay B General Conditions A brief description of the project, project name, Chapter-1 & Chapter-2 i nature, size, its importance to the region/state and the country shall be submitted In case the project involves diversion of forests No forest Land diversion land, guidelines under OM dated 20.03.2013 shall ii be followed and necessary action be taken accordingly Details of any litigation(s) pending against the No litigations are pending against project and/or any directions or orders passed by the project road iii any court of law/any statutory authority against the project to be detailed out.