Pair-Flowered Cymes in the Lamiales
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Newsletter 123 May 2012
TheTheThe Irish Garden Plant Society Newsletter Number 11123123 May 2012 The Annual General Meeting 2012 The Annual General Meeting will be held on Sat 12th May 10.00 a.m for 10.30 a.m., at Hillsborough Courthouse, The Square, Hillsborough, BT26 6AG. As always, it will be followed by a series of garden visits on Saturday & Sunday and a meal on Saturday evening. The meal will be held at 8:00pm in La Mon Hotel & Country Club, 41 Gransha Road, Comber, BT23 5RF. See the January 2012 newsletter for details of the gardens to be visited. If you haven’t already booked contact Patrick Quigley, 24 Areema Drive, Dunmurry, Belfast, BT17 0QG. tel: +44 (0) 7801 299263 [email protected] for further information. A.G. M. Agenda 1. Apologies 2. Minutes of AGM 2011 3. Matters arising 4. Chairman’s report 5. Treasurer’s report 6. Election of Committee Members 7. Any other business Front cover : Solanum crispum ‘Glasnevin’. Photograph : Pearse Rowe In this issue 2 Editorial 3 Southern Climbers for Northern Walls by John Joe Costin 11 Worth a Read by Paddy Tobin 14 The Palm House, a review by Mary Bradshaw 16 Cheers to Chiltern Seeds and to the Sole Survivor of Seed Project 1997/98 by Michael Kelleher 17 Seed Distribution Report 2011 and 2012 by Stephen Butler 19 Regional Reports 28 Spring at Kilmacurragh by Seamus O’Brien 37 Propagation of Arbutus by Kevin Line 41 Tulipa ‘Molly Bloom’ - an new Tulip for 2012 1 Editorial Spring time and new plants are synonymous, and as Christopher Lloyd said in Garden Flowers from Seed, seed sowing is “one of life’s big thrills”. -

Flowering Plants Eudicots Apiales, Gentianales (Except Rubiaceae)
Edited by K. Kubitzki Volume XV Flowering Plants Eudicots Apiales, Gentianales (except Rubiaceae) Joachim W. Kadereit · Volker Bittrich (Eds.) THE FAMILIES AND GENERA OF VASCULAR PLANTS Edited by K. Kubitzki For further volumes see list at the end of the book and: http://www.springer.com/series/1306 The Families and Genera of Vascular Plants Edited by K. Kubitzki Flowering Plants Á Eudicots XV Apiales, Gentianales (except Rubiaceae) Volume Editors: Joachim W. Kadereit • Volker Bittrich With 85 Figures Editors Joachim W. Kadereit Volker Bittrich Johannes Gutenberg Campinas Universita¨t Mainz Brazil Mainz Germany Series Editor Prof. Dr. Klaus Kubitzki Universita¨t Hamburg Biozentrum Klein-Flottbek und Botanischer Garten 22609 Hamburg Germany The Families and Genera of Vascular Plants ISBN 978-3-319-93604-8 ISBN 978-3-319-93605-5 (eBook) https://doi.org/10.1007/978-3-319-93605-5 Library of Congress Control Number: 2018961008 # Springer International Publishing AG, part of Springer Nature 2018 This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, and transmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology now known or hereafter developed. The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication does not imply, even in the absence of a specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use. -

Keene, Jeremy 11-21-13
A Reassessment of Monopyle (Gloxinieae:Gesneriaceae) A dissertation presented to the faculty of the College of Arts and Sciences of Ohio University In partial fulfillment of the requirements for the degree Doctor of Philosophy Jeremy L. Keene December 2013 © 2013 Jeremy L. Keene. All Rights Reserved. 2 This dissertation titled A Reassessment of Monopyle (Gloxinieae: Gesneriaceae) by JEREMY L. KEENE has been approved for the Department of Environmental and Plant Biology and the College of Arts and Sciences by Harvey E. Ballard, Jr. Associate Professor of Environmental and Plant Biology Robert Frank Dean, College of Arts and Sciences 3 ABSTRACT KEENE, JEREMY L., Ph.D., December 2013, Environmental and Plant Biology A Reassessment of Monopyle (Gloxinieae: Gesneriaceae) Director of Dissertation: Harvey E. Ballard, Jr. Monopyle Moritz ex Benth. is a genus of herbs or suffrutescent herbs distributed from Guatemala southward into northern South America. This assemblage of plants is characterized by the presence of anisophyllous leaves, uncinate trichomes, and campanulate flowers. The genus was last revised in 1945 by Conrad Morton. Since that time, few additional taxa have been recognized. It was comprised of 22 described species in 2011, with two infraspecific taxa. Also, there has been limited sampling of this genus for phylogenetic studies to assess the evolution of the genus. There were two primary foci of this research, a morphological characterization of the species within Monopyle and molecular analyses of the species within Monopyle and other closely related genera. A detailed morphological analysis was performed to determine the morphological differentiation between taxa. This analysis also confirmed which diagnostic characters identified in previous studies were valid and useful. -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -
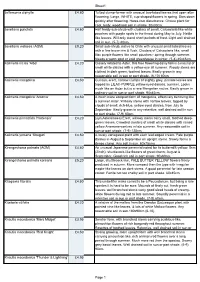
Sheet1 Jeffersonia Diphylla £4.80 Tufted Clump-Former with Unusual Two-Lobed Leaves That Open After Flowering
Sheet1 Jeffersonia diphylla £4.80 Tufted clump-former with unusual two-lobed leaves that open after flowering. Large, WHITE, cup-shaped flowers in spring. Dies down quickly after flowering. Hates root disturbance. Choice plant for cool, leafy, woodland soil in shade. 30x30cm. Jovellana punctata £4.60 Half hardy sub-shrub with clusters of small, Calceolaria like white pouches with purple spots in the throat during May to July. Nettle like leaves. Will only stand short periods of frost. Light well drained soil in sun. (5-7) 40cm. Jovellana violacea (AGM) £8.20 Small sub-shrub, native to Chile with unusual small lobed leaves with a fine brown rim & flush. Clusters of Calceolaria like, small lilac purple flowers like small pouches;- spring through summer. Needs a warm spot or cold greenhouse in winter. (5-8) 45x45cm. Kalimeris incisa 'Alba' £4.20 Closely related to Aster, this free flowering daisy forms a mound of small white daisies with a yellow eye all summer. Woody stems are clothed in dark green, toothed leaves. Easily grown in any reasonable soil in sun or part shade. (6-10) 60cm Kalimeris mongolica £4.50 Curious, erect, narrow clumps of slightly grey, pinnate leaves are topped by LILAC-PURPLE yellow-eyed daisies; summer. Looks much like an Aster but is a rare Mongolian native. Easily grown in ordinary soil in sun or part shade. 90x45cm. Kalimeris mongolica 'Antonia' £4.50 A much more compact form of mongolica, effectively behaving like a summer Aster. Willowy stems with narrow leaves, topped by clouds of small, rich-blue, yellow eyed daisies, from July to September. -

A New Application for Phylogenetic Marker Development Using Angiosperm Transcriptomes Author(S): Srikar Chamala, Nicolás García, Grant T
MarkerMiner 1.0: A New Application for Phylogenetic Marker Development Using Angiosperm Transcriptomes Author(s): Srikar Chamala, Nicolás García, Grant T. Godden, Vivek Krishnakumar, Ingrid E. Jordon- Thaden, Riet De Smet, W. Brad Barbazuk, Douglas E. Soltis, and Pamela S. Soltis Source: Applications in Plant Sciences, 3(4) Published By: Botanical Society of America DOI: http://dx.doi.org/10.3732/apps.1400115 URL: http://www.bioone.org/doi/full/10.3732/apps.1400115 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder. BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. ApApplicatitionsons Applications in Plant Sciences 2015 3 ( 4 ): 1400115 inin PlPlant ScienSciencesces S OFTWARE NOTE M ARKERMINER 1.0: A NEW APPLICATION FOR PHYLOGENETIC 1 MARKER DEVELOPMENT USING ANGIOSPERM TRANSCRIPTOMES S RIKAR C HAMALA 2,12 , N ICOLÁS G ARCÍA 2,3,4 * , GRANT T . G ODDEN 2,3,5 * , V IVEK K RISHNAKUMAR 6 , I NGRID E. -

Flower Food Tissues As Reward for Pollinating Birds
ISSN 0373-580 X Bol. Soc. Argent. Bot. 37 (1-2): 71 - 77. 2002 FLOWER FOOD TISSUES AS REWARD FOR POLLINATING BIRDS G. G. ROITMAN' 3, A. N. SÉRSIC23, A. A. COCUCCI2 and N. H. MONTALDO1 Summary: Nectar is the reward regularly offered by flowers to their pollinating birds. Studies on pollination by birds have focused on nectar as a reward while alternative reward systems have been poorly studied. Previous reviews have shown the presence of food tissues that serve as nutritive rewards to different classes of pollinators. However, in a recent revision of plant-bird interactions, the existence of a solid reward for birds has been disregarded. In South America at least three cases of this very particular bird-flower Interaction have been recently studied. In this paper we describe the syndrome features (flower morphology, reward properties, position of the pollination unit, flower color display and common visitors), that deviate from the typical ornithophilous syndrome, and its possible origin is discussed. Key words: Flower, food reward, pollination, birds, syndrome, review. Resumen: Tejidos alimenticios florales como recompensa para las aves polinizadoras. El néctar es la recompensa típica que las flores ofrecen a las aves polinizadoras. Los estudios sobre ornitofilia se han concentrado en el néctar como recompensa, mientras que otras retribuciones alternativas fueron esca¬ samente analizadas. Revisiones previas han mostrado que la producción de tejidos alimenticios puede servir como recompensa para diferentes polinizadores, y al menos tres casos de esta particular interacción se estudiaron últimamente en América del Sur. Sin embargo, una revisión reciente sobre interacciones planta-ave descarta la presencia de recompensas sólidas. -

Temporal and Spatial Origin of Gesneriaceae in the New World Inferred from Plastid DNA Sequences
bs_bs_banner Botanical Journal of the Linnean Society, 2013, 171, 61–79. With 3 figures Temporal and spatial origin of Gesneriaceae in the New World inferred from plastid DNA sequences MATHIEU PERRET1*, ALAIN CHAUTEMS1, ANDRÉA ONOFRE DE ARAUJO2 and NICOLAS SALAMIN3,4 1Conservatoire et Jardin botaniques de la Ville de Genève, Ch. de l’Impératrice 1, CH-1292 Chambésy, Switzerland 2Centro de Ciências Naturais e Humanas, Universidade Federal do ABC, Rua Santa Adélia, 166, Bairro Bangu, Santo André, Brazil 3Department of Ecology and Evolution, University of Lausanne, CH-1015 Lausanne, Switzerland 4Swiss Institute of Bioinformatics, Quartier Sorge, CH-1015 Lausanne, Switzerland Received 15 December 2011; revised 3 July 2012; accepted for publication 18 August 2012 Gesneriaceae are represented in the New World (NW) by a major clade (c. 1000 species) currently recognized as subfamily Gesnerioideae. Radiation of this group occurred in all biomes of tropical America and was accompanied by extensive phenotypic and ecological diversification. Here we performed phylogenetic analyses using DNA sequences from three plastid loci to reconstruct the evolutionary history of Gesnerioideae and to investigate its relationship with other lineages of Gesneriaceae and Lamiales. Our molecular data confirm the inclusion of the South Pacific Coronanthereae and the Old World (OW) monotypic genus Titanotrichum in Gesnerioideae and the sister-group relationship of this subfamily to the rest of the OW Gesneriaceae. Calceolariaceae and the NW genera Peltanthera and Sanango appeared successively sister to Gesneriaceae, whereas Cubitanthus, which has been previously assigned to Gesneriaceae, is shown to be related to Linderniaceae. Based on molecular dating and biogeographical reconstruction analyses, we suggest that ancestors of Gesneriaceae originated in South America during the Late Cretaceous. -

Hidden in Plain Sight—A New Species of Lichen Strigula Oleistrata March 2020 (Strigulaceae) from New Zealand
TRILEPIDEA Newsletter of the New Zealand Plant Conservation Network NO. 196 Hidden in Plain sight—a new species of lichen Strigula oleistrata March 2020 (Strigulaceae) from New Zealand. Deadline for next issue: Marley Ford ([email protected]); Dan J. Blanchon ([email protected]), Friday 19 April 2020 School of Environmental & Animal Sciences, Unitec Institute of Technology, Auckland; SUBMIT AN ARTICLE Peter J. de Lange ([email protected]), School of Environmental & Animal TO THE NEWSLETTER Sciences, Unitec Institute of Technology, Auckland Contributions are welcome New Zealand has a surprising diversity of Strigula. 25 species of the genus are found to the newsletter at any here out of the c.70 species recognised; representing over a third of the known species time. The closing date for articles for each issue is (Galloway 2007, Lücking 2008, Hyde et al. 2013). Most of these species are foliicolous, approximately the 15th of meaning that they live on the surfaces of leaves. However, a few species do colonise each month. rocks and bark (Galloway 2007). Articles may be edited and used in the newsletter and/ Between 2016 and 2017 the senior author undertook a third year, level 7 School of or on the website news page. Environmental & Animal Sciences self-directed paper studying Strigula novae- The Network will publish zelandiae at the Unitec Institute of Technology Herbarium (UNITEC). Strigula novae- almost any article about zelandiae is a foliicolous species that is sometimes known as ‘silver paint lichen’, because plants and plant conservation with a particular focus on the when dead the thallus imparts a silvery patterning on the leaves it has colonised. -

Biological Resources Assessment the Ranch ±530- Acre Study Area City of Rancho Cordova, California
Biological Resources Assessment The Ranch ±530- Acre Study Area City of Rancho Cordova, California Prepared for: K. Hovnanian Homes October 13, 2017 Prepared by: © 2017 TABLE OF CONTENTS 1.0 Introduction ......................................................................................................................... 1 1.1. Project Description ........................................................................................................... 1 2.0 Regulatory Framework ........................................................................................................ 2 2.1. Federal Regulations .......................................................................................................... 2 2.1.1. Federal Endangered Species Act ............................................................................... 2 2.1.2. Migratory Bird Treaty Act ......................................................................................... 2 2.1.3. The Bald and Golden Eagle Protection Act ............................................................... 2 2.2. State Jurisdiction .............................................................................................................. 3 2.2.1. California Endangered Species Act ........................................................................... 3 2.2.2. California Department of Fish and Game Codes ...................................................... 3 2.2.3. Native Plant Protection Act ..................................................................................... -

Towards Resolving Lamiales Relationships
Schäferhoff et al. BMC Evolutionary Biology 2010, 10:352 http://www.biomedcentral.com/1471-2148/10/352 RESEARCH ARTICLE Open Access Towards resolving Lamiales relationships: insights from rapidly evolving chloroplast sequences Bastian Schäferhoff1*, Andreas Fleischmann2, Eberhard Fischer3, Dirk C Albach4, Thomas Borsch5, Günther Heubl2, Kai F Müller1 Abstract Background: In the large angiosperm order Lamiales, a diverse array of highly specialized life strategies such as carnivory, parasitism, epiphytism, and desiccation tolerance occur, and some lineages possess drastically accelerated DNA substitutional rates or miniaturized genomes. However, understanding the evolution of these phenomena in the order, and clarifying borders of and relationships among lamialean families, has been hindered by largely unresolved trees in the past. Results: Our analysis of the rapidly evolving trnK/matK, trnL-F and rps16 chloroplast regions enabled us to infer more precise phylogenetic hypotheses for the Lamiales. Relationships among the nine first-branching families in the Lamiales tree are now resolved with very strong support. Subsequent to Plocospermataceae, a clade consisting of Carlemanniaceae plus Oleaceae branches, followed by Tetrachondraceae and a newly inferred clade composed of Gesneriaceae plus Calceolariaceae, which is also supported by morphological characters. Plantaginaceae (incl. Gratioleae) and Scrophulariaceae are well separated in the backbone grade; Lamiaceae and Verbenaceae appear in distant clades, while the recently described Linderniaceae are confirmed to be monophyletic and in an isolated position. Conclusions: Confidence about deep nodes of the Lamiales tree is an important step towards understanding the evolutionary diversification of a major clade of flowering plants. The degree of resolution obtained here now provides a first opportunity to discuss the evolution of morphological and biochemical traits in Lamiales. -

Evolutionary History of Floral Key Innovations in Angiosperms Elisabeth Reyes
Evolutionary history of floral key innovations in angiosperms Elisabeth Reyes To cite this version: Elisabeth Reyes. Evolutionary history of floral key innovations in angiosperms. Botanics. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS489. tel-01443353 HAL Id: tel-01443353 https://tel.archives-ouvertes.fr/tel-01443353 Submitted on 23 Jan 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS489 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de Doctorat : Biologie Par Mme Elisabeth Reyes Evolutionary history of floral key innovations in angiosperms Thèse présentée et soutenue à Orsay, le 13 décembre 2016 : Composition du Jury : M. Ronse de Craene, Louis Directeur de recherche aux Jardins Rapporteur Botaniques Royaux d’Édimbourg M. Forest, Félix Directeur de recherche aux Jardins Rapporteur Botaniques Royaux de Kew Mme. Damerval, Catherine Directrice de recherche au Moulon Président du jury M. Lowry, Porter Curateur en chef aux Jardins Examinateur Botaniques du Missouri M. Haevermans, Thomas Maître de conférences au MNHN Examinateur Mme. Nadot, Sophie Professeur à l’Université Paris-Sud Directeur de thèse M.