Intraguild Predation and Interspecific Killing As Structuring Forces of Carnivoran Communities in South America
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fur Trade and the Biotic Homogenization of Subpolar Ecosystems
Chapter 14 Fur Trade and the Biotic Homogenization of Subpolar Ecosystems Ramiro D. Crego, Ricardo Rozzi, and Jaime E. Jiménez Abstract At the southern end of the Americas exist one of the last pristine ecosys- tems in the world, the sub-Antarctic Magellanic forests ecoregion, protected by the Cape Horn Biosphere Reserve (CHBR). Despite its remote location, the CHBR has been subject to the growing infuences of globalization, a process that has driven cultural, biotic, and economic transformations in the region since the mid-twentieth century. One of the most important threats to these unique ecosystems is the increase of biological invasions. Motivated by the expanding fur industry that responded to the globalization process, American beavers (Castor canadensis), muskrats (Ondatra zibethicus), and American minks (Neovison vison) were introduced, inde- pendently, to the southern tip of South America. Research has shown that these three North American species have reassembled their native interactions to affect negatively the invaded ecosystems of the CHBR. Beavers affect river fow and R. D. Crego (*) Department of Biological Sciences, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad, Santiago, Chile Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA R. Rozzi Department of Philosophy and Religion and Department of Biological Sciences, University of North Texas, Denton, TX, USA Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad and Universidad de Magallanes, Punta Arenas, Chile J. E. Jiménez Department of Biological Sciences, University of North Texas, Denton, TX, USA Instituto de Ecología y Biodiversidad, Santiago, Chile Sub-Antarctic Biocultural Conservation Program, University of North Texas, Denton, TX, USA Department of Philosophy and Religion, University of North Texas, Denton, TX, USA Universidad de Magallanes, Punta Arenas, Chile © Springer Nature Switzerland AG 2018 233 R. -

Phylogeographic and Diversification Patterns of the White-Nosed Coati
Molecular Phylogenetics and Evolution 131 (2019) 149–163 Contents lists available at ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev Phylogeographic and diversification patterns of the white-nosed coati (Nasua narica): Evidence for south-to-north colonization of North America T ⁎ Sergio F. Nigenda-Moralesa, , Matthew E. Gompperb, David Valenzuela-Galvánc, Anna R. Layd, Karen M. Kapheime, Christine Hassf, Susan D. Booth-Binczikg, Gerald A. Binczikh, Ben T. Hirschi, Maureen McColginj, John L. Koprowskik, Katherine McFaddenl,1, Robert K. Waynea, ⁎ Klaus-Peter Koepflim,n, a Department of Ecology & Evolutionary Biology, University of California, Los Angeles, Los Angeles, CA 90095, USA b School of Natural Resources, University of Missouri, Columbia, MO 65211, USA c Departamento de Ecología Evolutiva, Centro de Investigación en Biodiversidad y Conservación, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos 62209, Mexico d Department of Pathology and Laboratory Medicine, David Geffen School of Medicine, University of California, Los Angeles, Los Angeles, CA 90095, USA e Department of Biology, Utah State University, Logan, UT 84322, USA f Wild Mountain Echoes, Vail, AZ 85641, USA g New York State Department of Environmental Conservation, Albany, NY 12233, USA h Amsterdam, New York 12010, USA i Zoology and Ecology, College of Science and Engineering, James Cook University, Townsville, QLD 4811, Australia j Department of Biological Sciences, Purdue University, West Lafayette, IN 47907, USA k School of Natural Resources and the Environment, The University of Arizona, Tucson, AZ 85721, USA l College of Agriculture, Forestry and Life Sciences, Clemson University, Clemson, SC 29634, USA m Smithsonian Conservation Biology Institute, National Zoological Park, Washington, D.C. -
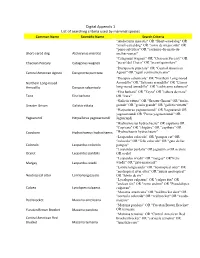
Digital Appendix 1 List of Searching Criteria Used by Mammal Species
Digital Appendix 1 List of searching criteria used by mammal species Common Name Scientific Name Search Criteria "Atelocynus microtis" OR "Short-eared dog" OR "small-eared dog" OR "zorro de oreja corta" OR "perro selvático" OR "cachorro-do-mato-de- Short-eared dog Atelocynus microtis orelhas-curtas" "Catagonus wagneri" OR "Chacoan Peccary" OR Chacoan Peccary Catagonus wagneri "pecarí del Chaco" OR "pecarí quimilero" “Dasyprocta punctate” OR "Central American Central American Agouti Dasyprocta punctata Agouti" OR "agutí centroamericano" “Dasypus sabanicola” OR "Northern Long-nosed Northern Long-nosed Armadillo" OR "Savanna armadillo" OR "Llanos Armadillo Dasypus sabanicola long-nosed armadillo" OR "cachicamo sabanero" “Eira barbara” OR "Tayra" OR "cabeza de mate" Taira Eira barbara OR "irara" “Galictis vittata” OR "Greater Grison" OR "furão- Greater Grison Galictis vittata grande" OR "grisón grande" OR "galictis vittatta" “Herpailurus yagouaroundi” OR Yaguarundi OR yagouaroundi OR "Puma yagouaroundi" OR Yaguarundi Herpailurus yagouaroundi Jaguarundi "Hydrochoerus hydrochaeris" OR capybara OR "Capivara" OR "chigüire" OR "capibara" OR Capybara Hydrochoerus hydrochaeris "Hydrochaeris hydrochaeris" “Leopardus colocolo” OR "pampas cat" OR "colocolo" OR "felis colocolo" OR "gato de las Colocolo Leopardus colocolo pampas" "Leopardus pardalis" OR jaguatirica OR ocelote Ocelot Leopardus pardalis OR ocelot “Leopardus wiedii” OR "margay" OR"Felis Margay Leopardus wiedii wiedii" OR "gato-maracajá" “Lontra longicaudis” OR "neotropical otter" OR "neotropical -

APROXIMACIÓN AL LENGUAJE REPERTORIAL DE LAS ESTEREOTIPIAS EN MAMIFEROS CARNIVOROS JAGUAR (Panthera Onca), GRISÓN (Galictis
APROXIMACIÓN AL LENGUAJE REPERTORIAL DE LAS ESTEREOTIPIAS EN MAMIFEROS CARNIVOROS JAGUAR (Panthera onca), GRISÓN (Galictis vittata) Y OCELOTE (Leopardus pardalis) DEL ZOOLOGICO GUATIKA, BOYACA – TIBASOSA. Laura Inés Monsalve Zuleta CÓDIGO:2014101101 Trabajo de investigación presentado como requisito parcial para optar al título de: Médica veterinaria especialista en Medicina Homeopática veterinaria Director (a): Néstor Alberto Calderón Maldonado Médico Veterinario – Universidad de la Salle Certificado en Medicina Veterinaria Homeopática – FICH “Luis G. Páez” Especialista en Bioética – Universidad El Bosque Fundación Universitaria Escuela Colombiana de Medicina Homeopática “Luis G. Páez”. Programa de Medicina Homeopática Veterinaria Bogotá, Colombia 2016 2 DEDICATORIA A mi familia, amigos y doctores que me apoyaron en esta etapa de mi vida tanto profesional como personal. 3 AGRADECIMIENTOS Agradezco a mi familia, amigos y Doctores que me ayudaron a culminar esta etapa de mi vida. 4 TABLA DE CONTENIDO: 1. RESUMEN ...................................................................................................................................... 7 2. INTRODUCCIÓN ........................................................................................................................... 8 3. JUSTIFICACIÓN ........................................................................................................................... 9 4. ESTADO DEL ARTE ................................................................................................................. -

Proceedings of the United States National Museum
THE GENERA AND SUBGENERA OF RACCOONS AND THEIR ALLIES. By N. HOLLISTER, Assistant Curator, Division of Mammals, United States National Museum. The family Procyonidae offers a remarkable instance of a group of mammal genera in which the differentiation in the structure of the teeth throughout the series is nearly uniform in degree from genus to genus, and strictly definite in direction from one extreme to the other. Tliis condition is particularly interesting because of the wide difference between the teeth of genera from the extremes of the line, and because the widely diverse cranial and external charac- ters show no special tendency toward serial grouping. EUminating the very aberrant Bassariscus, which clearly does not belong in this series of genera, the superspecific groups remaining within the Procyonidge, when based chiefly on dental characteristics, fall into as weU-ordered a sequence of steps, connecting the Old-World Ailurus with the American Potos, as it would seem possible to find among living animals. (Plate 39.) The one seemingly aberrant cranial feature in Ailurus,^ the presence of the aUsphenoid canal, can not in tliis case be considered a character of family or subfamily importance. The alisphenoid canal in other famiUes of carnivores is known to be absent or present in different genera, in individuals of the same species, or even on the right and left side of the same individual. Of the American groups, Euprocyon is the nearest approach to Ailurus, but if the genera within the family are to be kept of fairly uniform value and degree of differen- tiation, Euprocyon is surely not more than a subgenus of Procyon. -

Predicting Greater Grison Galictis Vittata Presence from Scarce Records in the Department of Cordoba, Colombia
ORIGINAL CONTRIBUTION Predicting Greater Grison Galictis vittata presence from scarce records in the department of Cordoba, Colombia José F. GONZÁLEZ-MAYA1*, Julio J. CHACÓN PACHECO2, Javier RACERO-CASARRUBIA2, Erika HUMANEZ-LÓPEZ2 & Andrés ARIAS-ALZATE3 1. Proyecto de Conservación de Aguas y Tierras, ProCAT Colombia/Internacional, Calle 97ª # 10-62, Of. 202, Bogotá, Abstract. Colombia. The Greater Grison, Galictis vittata, is a poorly known species in Colombia. Throughout 2. Grupo de Investigación its range major knowledge gaps exist regarding its ecology and conservation. To compile Biodiversidad Unicórdoba, and analyse information about the species´ distribution records in the department of Universidad de Córdoba, Cordoba, Colombia and assess its presence probability according to landscape attributes, Montería, Córdoba, Colombia. we conducted a literature review of all wildlife studies in the region and compiled all possible direct presence records of the species in the department. We generated random 3.Grupo de Mastozoología, location points and characterized each distribution and random location by their distance to landscape attributes and land-cover type and modelled landscape presence using a Instituto de Biología, Multiple Logistic Regression approach. We found 33 records of the species in Cordoba Universidad de Antioquía. with most of the records distributed in the subregion of Alto Sinú (36%). Higher presence Medellín, Colombia. probabilities are localized in areas near forests mostly in the southern parts of the department, showing the species is still related with the largest forest blocks. Grisons Correspondence: appears to potentially tolerate some levels of disturbance but is still dependent to forest. The influence of natural habitats and abundance across the department and other areas José F. -

Controlled Animals
Environment and Sustainable Resource Development Fish and Wildlife Policy Division Controlled Animals Wildlife Regulation, Schedule 5, Part 1-4: Controlled Animals Subject to the Wildlife Act, a person must not be in possession of a wildlife or controlled animal unless authorized by a permit to do so, the animal was lawfully acquired, was lawfully exported from a jurisdiction outside of Alberta and was lawfully imported into Alberta. NOTES: 1 Animals listed in this Schedule, as a general rule, are described in the left hand column by reference to common or descriptive names and in the right hand column by reference to scientific names. But, in the event of any conflict as to the kind of animals that are listed, a scientific name in the right hand column prevails over the corresponding common or descriptive name in the left hand column. 2 Also included in this Schedule is any animal that is the hybrid offspring resulting from the crossing, whether before or after the commencement of this Schedule, of 2 animals at least one of which is or was an animal of a kind that is a controlled animal by virtue of this Schedule. 3 This Schedule excludes all wildlife animals, and therefore if a wildlife animal would, but for this Note, be included in this Schedule, it is hereby excluded from being a controlled animal. Part 1 Mammals (Class Mammalia) 1. AMERICAN OPOSSUMS (Family Didelphidae) Virginia Opossum Didelphis virginiana 2. SHREWS (Family Soricidae) Long-tailed Shrews Genus Sorex Arboreal Brown-toothed Shrew Episoriculus macrurus North American Least Shrew Cryptotis parva Old World Water Shrews Genus Neomys Ussuri White-toothed Shrew Crocidura lasiura Greater White-toothed Shrew Crocidura russula Siberian Shrew Crocidura sibirica Piebald Shrew Diplomesodon pulchellum 3. -

Abundance Changes and Activity Flexibility of the Oncilla, Leopardus Tigrinus (Carnivora: Felidae), Appear to Reflect Avoidance of Conflict
ZOOLOGIA 29 (2): 115–120, April, 2012 doi: 10.1590/S1984-46702012000200003 Abundance changes and activity flexibility of the oncilla, Leopardus tigrinus (Carnivora: Felidae), appear to reflect avoidance of conflict Luiz Gustavo R. Oliveira-Santos1, 6, Maurício E. Graipel2, Marcos A. Tortato3, Carlos A. Zucco1, Nilton C. Cáceres4 & Fernando V. B. Goulart5 1 Laboratório de Ecologia e Conservação de Populações, Departamento de Ecologia, Universidade Federal do Rio de Janeiro. Caixa Postal 68020, 21941-590 Rio de Janeiro, RJ, Brazil. 2 Departamento de Ecologia e Zoologia, Universidade Federal de Santa Catarina. 88040-970 Florianópolis, SC, Brazil. 3 Programa de Pós-graduação em Ecologia e Conservação, Setor de Ciências Biológicas, Universidade Federal do Paraná. 81531-980, Curitiba, PR, Brazil. 4 Departamento de Biologia, Universidade Federal de Santa Maria. 97105-900 Santa Maria, RS, Brazil. 5 Programa de Pós-graduação em Ecologia e Conservação, Universidade Federal do Mato Grosso do Sul. 79070-900 Campo Grande, MS, Brazil. 6 Corresponding Author. Email: [email protected] ABSTRACT. We investigated the density and activity of the oncilla, Leopardus tigrinus (Schreber, 1775), a threatened small cat, in the Brazilian Atlantic Forest, using camera-trap data. We described differences in the activity of individuals occurring alone or in sympatry with larger cats. Oncilla presented low densities (7-13 ind./100 km²) and high flexibility in its activity. The oncillas were primarily nocturnal in the absence of other larger cat species – margay, ocelot and puma – but became more diurnal, with a cathemeral activity pattern, when the other cats were present. Oncilla is likely to be in a subordinate position in interactions with larger cats and changes its activity to decrease the chances for interspecific encounters. -

Galictis Cuja Molina, 1782) As Host of Dioctophyme Renale Goeze, 1782 Furão Pequeno (Galictis Cuja Molina, 1782) Como Hospedeiro De Dioctophyme Renale Goeze, 1782
ANIMAL PARASITOLOGY / SCIENTIFIC COMMUNICATION DOI: 10.1590/1808-1657000312016 Lesser Grison (Galictis cuja Molina, 1782) as host of Dioctophyme renale Goeze, 1782 Furão Pequeno (Galictis cuja Molina, 1782) como hospedeiro de Dioctophyme renale Goeze, 1782 Daniela Pedrassani1*, Mayana Worm1, Jéssica Drechmer1, Margareth Cristina Iazzetti Santos1 ABSTRACT: The Dioctophyme renale is a helminth parasite RESUMO: O Dioctophyme renale é um helminto parasita renal of the kidney usually seen in domestic and wild carnivores and observado normalmente em carnívoros domésticos e silvestres e rarely in human beings. This is a report about the parasitism excepcionalmente em seres humanos. Relata-se o parasitismo por D. of D. renale found in the kidney of two roadkill lesser grisons renale em rim de dois furões pequenos (Galictis cuja) encontrados (Galictis cuja) in the North of the state of Santa Catarina, mortos por atropelamento no Norte do estado de Santa Catarina, Brazil. The report of this parasitism in this species is important Brasil. Relatar esse parasitismo nessa espécie é importante, para to complement the records about this native carnivore as a que se possam somar dados relativos a participação deste carnívoro contributor in the epidemiologic chain while host/disseminator nativo na cadeia epidemiológica como hospedeiro/ veiculador desse of this helminth with zoonotic potential. helminto com potencial zoonótico. KEYWORDS: Dioctophyma; wild animal; mustelids; roadkill; PALAVRAS-CHAVE: Dioctophyma; animal silvestre; mustelí- kidney parasitism. deo; atropelamento em rodovia; parasitismo renal. 1Universidade do Contestado (UnC) – Canoinhas (SC), Brazil. *Corresponding author: [email protected] Received on: 04/22/2016. Accepted on: 09/12/2017 Arq. Inst. Biol., v.84, 1-4, e0312016, 2017 1 D. -

Additions to Mammals Killed by Motor Vehicles on Vía Del Escobero, Envigado
Revista EIA, ISSN 1794-1237 / Year XI / Volume 11 / Issue N.22 / July-December 2014 / pp. 137-142 Technical-scientific biannual publication / Escuela de Ingeniería de Antioquia —EIA—, Envigado (Colombia) ADDITIONS TO MAMMALS KILLED BY MOTOR VEHICLES ON VÍA DEL ESCOBERO, ENVIGADO CARLOS A. DELGADO-V. 1 ABSTRACT Vertebrate road kills are a generalized problem around the world, but they are scarcely documented on Colombian highways, especially in peri-urban areas. This study describes the highway mortality of mammals in a six year-period (2008-2013) on Vía del Escobero (Envigado, Antioquia, Colombia). Mammal groups that presented the highest mortality rates were marsupials (54.3%), carnivores (25.7%), and rodents (17.5%). Although there was a lower diversity and fewer individual fatalities than in previous years for the same highway, this study reports a greater number of fatalities for some endangered and unknown species such as Leopardus tigrinus, Puma yag- ouaroundi, and Bassaricyon neblina. This study also added three new species to the road kill list on this highway. KEYWORDS: Road ecology, urban ecology, Medellín, Valle de Aburrá, Valle de San Nicolás ADICIONES AL ATROPELLAMIENTO VEHICULAR DE MAMÍFEROS EN LA VÍA DE EL ESCOBERO, ENVIGADO (ANTIOQUIA), COLOMBIA RESUMEN El atropellamiento vehicular de fauna es un problema generalizado alrededor del Mundo pero escasamente estudiado en las carreteras colombianas, especialmente en áreas periurbanas. Aquí se describe la mortalidad de mamíferos entre los años 2008 a 2013 en la vía El Escobero (Envigado, Antioquia). Los mamíferos más atropella- dos en este periodo fueron los marsupiales (54.3 %), los carnívoros (25.7 %) y los roedores (17.5 %). -

Home Range and Spatial Organization by the Hoary Fox Lycalopex Vetulus (Mammalia: Carnivora: Canidae): Response to Social Disruption of Two Neighboring Pairs
OPEN ACCESS The Journal of Threatened Taxa is dedicated to building evidence for conservaton globally by publishing peer-reviewed artcles online every month at a reasonably rapid rate at www.threatenedtaxa.org. All artcles published in JoTT are registered under Creatve Commons Atributon 4.0 Internatonal License unless otherwise mentoned. JoTT allows unrestricted use of artcles in any medium, reproducton, and distributon by providing adequate credit to the authors and the source of publicaton. Journal of Threatened Taxa Building evidence for conservaton globally www.threatenedtaxa.org ISSN 0974-7907 (Online) | ISSN 0974-7893 (Print) Communication Home range and spatial organization by the Hoary Fox Lycalopex vetulus (Mammalia: Carnivora: Canidae): response to social disruption of two neighboring pairs Julio C. Dalponte, Herson S. Lima, Stuart Klorfne & Nelton C. da Luz 26 May 2018 | Vol. 10 | No. 6 | Pages: 11703–11709 10.11609/jot.3082.10.6.11703-11709 For Focus, Scope, Aims, Policies and Guidelines visit htp://threatenedtaxa.org/index.php/JoTT/about/editorialPolicies#custom-0 For Artcle Submission Guidelines visit htp://threatenedtaxa.org/index.php/JoTT/about/submissions#onlineSubmissions For Policies against Scientfc Misconduct visit htp://threatenedtaxa.org/index.php/JoTT/about/editorialPolicies#custom-2 For reprints contact <[email protected]> Publisher & Host Partners Member Threatened Taxa Journal of Threatened Taxa | www.threatenedtaxa.org | 26 May 2018 | 10(6): 11703–11709 Home range and spatial organization by the Hoary Fox Lycalopex vetulus (Mammalia: Carnivora: Canidae): response Communication to social disruption of two neighboring pairs ISSN 0974-7907 (Online) ISSN 0974-7893 (Print) Julio C. Dalponte 1, Herson S. Lima 2, Stuart Klorfne 3 & Nelton C. -

Temporal Changes in the Diet of Two Sympatric Carnivorous Mammals in a Protected Area of South–Central Chile Affected by a Mixed–Severity Forest Fire
Animal Biodiversity and Conservation 43.2 (2020) 177 Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire A. H. Zúñiga, J. R. Rau, V. Fuenzalida, A. Fuentes–Ramírez Zúñiga, A. H., Rau, J. R., Fuenzalida, V., Fuentes–Ramírez, A., 2020. Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire. Animal Biodiversity and Conservation, 43.2: 177–186, Doi: https://doi.org/10.32800/abc.2020.43.0177 Abstract Temporal changes in the diet of two sympatric carnivorous mammals in a protected area of south–central Chile affected by a mixed–severity forest fire. Fire is a significant disruptive agent in various ecosystems around the world. It can affect the availability of resources in a given area, modulating the interaction between competing species. We studied the diet of the culpeo fox (Lycalopex culpaeus) and cougar (Puma concolor) for two consecutive years in a protected area of southern–central Chile which was affected by a wildfire. Significant differences were observed in the dietary pattern between the two species, showing their trophic segregation. In the two years of the study, the predominant prey for cougar was an exotic species, the European hare (Lepus europaeus), implying a simplification of its trophic spectrum with respect to that reported in other latitudes. The ecological consequences related to this scenario are discussed. Key words: Dietary overlap, Predation, Post–fire dynamics, Microhabitat, Rodent cycles, Selectivity Resumen Cambios temporales en la dieta de dos mamíferos carnívoros simpátridas en una zona protegida del centro y el sur de Chile afectada por un incendio forestal de intensidad desigual.