The Evolutionary Significance of the Wajak Skulls
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Quaternary International 603 (2021) 40–63
Quaternary International 603 (2021) 40–63 Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint Taxonomy, taphonomy and chronology of the Pleistocene faunal assemblage at Ngalau Gupin cave, Sumatra Holly E. Smith a,*, Gilbert J. Price b, Mathieu Duval c,a, Kira Westaway d, Jahdi Zaim e, Yan Rizal e, Aswan e, Mika Rizki Puspaningrum e, Agus Trihascaryo e, Mathew Stewart f, Julien Louys a a Australian Research Centre for Human Evolution, Environmental Futures Research Institute, Griffith University, Nathan, Queensland, 4111, Australia b School of Earth and Environmental Sciences, The University of Queensland, St Lucia, Queensland, 4072, Australia c Centro Nacional de Investigacion´ Sobre la Evolucion´ Humana (CENIEH), Burgos, 09002, Spain d Department of Earth and Environmental Sciences, Macquarie University, Sydney, New South Wales, Australia e Geology Study Program, Institut Teknologi Bandung, Jawa Barat, 40132, Indonesia f Extreme Events Research Group, Max Planck Institutes for Chemical Ecology, the Science of Human History, and Biogeochemistry, Jena, Germany ARTICLE INFO ABSTRACT Keywords: Ngalau Gupin is a broad karstic cave system in the Padang Highlands of western Sumatra, Indonesia. Abundant Taxonomy fossils, consisting of mostly isolated teeth from small-to large-sized animals, were recovered from breccias Taphonomy cemented on the cave walls and unconsolidated sediments on the cave floor.Two loci on the walls and floorsof Cave Ngalau Gupin, named NG-A and NG-B respectively, are studied. We determine that NG-B most likely formed as a Pleistocene result of the erosion and redeposition of material from NG-A. The collection reveals a rich, diverse Pleistocene Southeast Asia Hexaprotodon faunal assemblage (Proboscidea, Primates, Rodentia, Artiodactyla, Perissodactyla, Carnivora) largely analogous ESR and U-series dating to extant fauna in the modern rainforests of Sumatra. -

Endangered Species (Protection, Conser Va Tion and Regulation of Trade)
ENDANGERED SPECIES (PROTECTION, CONSER VA TION AND REGULATION OF TRADE) THE ENDANGERED SPECIES (PROTECTION, CONSERVATION AND REGULATION OF TRADE) ACT ARRANGEMENT OF SECTIONS Preliminary Short title. Interpretation. Objects of Act. Saving of other laws. Exemptions, etc., relating to trade. Amendment of Schedules. Approved management programmes. Approval of scientific institution. Inter-scientific institution transfer. Breeding in captivity. Artificial propagation. Export of personal or household effects. PART I. Administration Designahem of Mana~mentand establishment of Scientific Authority. Policy directions. Functions of Management Authority. Functions of Scientific Authority. Scientific reports. PART II. Restriction on wade in endangered species 18. Restriction on trade in endangered species. 2 ENDANGERED SPECIES (PROTECTION, CONSERVATION AND REGULA TION OF TRADE) Regulation of trade in species spec fled in the First, Second, Third and Fourth Schedules Application to trade in endangered specimen of species specified in First, Second, Third and Fourth Schedule. Export of specimens of species specified in First Schedule. Importation of specimens of species specified in First Schedule. Re-export of specimens of species specified in First Schedule. Introduction from the sea certificate for specimens of species specified in First Schedule. Export of specimens of species specified in Second Schedule. Import of specimens of species specified in Second Schedule. Re-export of specimens of species specified in Second Schedule. Introduction from the sea of specimens of species specified in Second Schedule. Export of specimens of species specified in Third Schedule. Import of specimens of species specified in Third Schedule. Re-export of specimens of species specified in Third Schedule. Export of specimens specified in Fourth Schedule. PART 111. -

Pocket Atlas of Human Anatomy 4Th Edition
I Pocket Atlas of Human Anatomy 4th edition Feneis, Pocket Atlas of Human Anatomy © 2000 Thieme All rights reserved. Usage subject to terms and conditions of license. III Pocket Atlas of Human Anatomy Based on the International Nomenclature Heinz Feneis Wolfgang Dauber Professor Professor Formerly Institute of Anatomy Institute of Anatomy University of Tübingen University of Tübingen Tübingen, Germany Tübingen, Germany Fourth edition, fully revised 800 illustrations by Gerhard Spitzer Thieme Stuttgart · New York 2000 Feneis, Pocket Atlas of Human Anatomy © 2000 Thieme All rights reserved. Usage subject to terms and conditions of license. IV Library of Congress Cataloging-in-Publication Data is available from the publisher. 1st German edition 1967 2nd Japanese edition 1983 7th German edition 1993 2nd German edition 1970 1st Dutch edition 1984 2nd Dutch edition 1993 1st Italian edition 1970 2nd Swedish edition 1984 2nd Greek edition 1994 3rd German edition 1972 2nd English edition 1985 3rd English edition 1994 1st Polish edition 1973 2nd Polish edition 1986 3rd Spanish edition 1994 4th German edition 1974 1st French edition 1986 3rd Danish edition 1995 1st Spanish edition 1974 2nd Polish edition 1986 1st Russian edition 1996 1st Japanese edition 1974 6th German edition 1988 2nd Czech edition 1996 1st Portuguese edition 1976 2nd Italian edition 1989 3rd Swedish edition 1996 1st English edition 1976 2nd Spanish edition 1989 2nd Turkish edition 1997 1st Danish edition 1977 1st Turkish edition 1990 8th German edition 1998 1st Swedish edition 1979 1st Greek edition 1991 1st Indonesian edition 1998 1st Czech edition 1981 1st Chinese edition 1991 1st Basque edition 1998 5th German edition 1982 1st Icelandic edition 1992 3rd Dutch edtion 1999 2nd Danish edition 1983 3rd Polish edition 1992 4th Spanish edition 2000 This book is an authorized and revised translation of the 8th German edition published and copy- righted 1998 by Georg Thieme Verlag, Stuttgart, Germany. -

Morfofunctional Structure of the Skull
N.L. Svintsytska V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 Ministry of Public Health of Ukraine Public Institution «Central Methodological Office for Higher Medical Education of MPH of Ukraine» Higher State Educational Establishment of Ukraine «Ukranian Medical Stomatological Academy» N.L. Svintsytska, V.H. Hryn Morfofunctional structure of the skull Study guide Poltava 2016 2 LBC 28.706 UDC 611.714/716 S 24 «Recommended by the Ministry of Health of Ukraine as textbook for English- speaking students of higher educational institutions of the MPH of Ukraine» (minutes of the meeting of the Commission for the organization of training and methodical literature for the persons enrolled in higher medical (pharmaceutical) educational establishments of postgraduate education MPH of Ukraine, from 02.06.2016 №2). Letter of the MPH of Ukraine of 11.07.2016 № 08.01-30/17321 Composed by: N.L. Svintsytska, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor V.H. Hryn, Associate Professor at the Department of Human Anatomy of Higher State Educational Establishment of Ukraine «Ukrainian Medical Stomatological Academy», PhD in Medicine, Associate Professor This textbook is intended for undergraduate, postgraduate students and continuing education of health care professionals in a variety of clinical disciplines (medicine, pediatrics, dentistry) as it includes the basic concepts of human anatomy of the skull in adults and newborns. Rewiewed by: O.M. Slobodian, Head of the Department of Anatomy, Topographic Anatomy and Operative Surgery of Higher State Educational Establishment of Ukraine «Bukovinian State Medical University», Doctor of Medical Sciences, Professor M.V. -

The First Occurrence of Eurygnathohippus Van Hoepen, 1930 (Mammalia, Perissodactyla, Equidae) Outside Africa and Its Biogeograph
TO L O N O G E I L C A A P I ' T A A T L E I I A Bollettino della Società Paleontologica Italiana, 58 (2), 2019, 171-179. Modena C N O A S S. P. I. The frst occurrence of Eurygnathohippus Van Hoepen, 1930 (Mammalia, Perissodactyla, Equidae) outside Africa and its biogeographic signifcance Advait Mahesh Jukar, Boyang Sun, Avinash C. Nanda & Raymond L. Bernor A.M. Jukar, Department of Paleobiology, National Museum of Natural History, Smithsonian Institution, Washington DC 20013, USA; [email protected] B. Sun, Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology Paleoanthropology, Chinese Academy of Sciences, Beijing 100044, China; University of Chinese Academy of Sciences, Beijing 100039, China; College of Medicine, Department of Anatomy, Laboratory of Evolutionary Biology, Howard University, Washington D.C. 20059, USA; [email protected] A.C. Nanda, Wadia Institute of Himalayan Geology, Dehra Dun 248001, India; [email protected] R.L. Bernor, College of Medicine, Department of Anatomy, Laboratory of Evolutionary Biology, Howard University, Washington D.C. 20059, USA; [email protected] KEY WORDS - South Asia, Pliocene, Biogeography, Dispersal, Siwalik, Hipparionine horses. ABSTRACT - The Pliocene fossil record of hipparionine horses in the Indian Subcontinent is poorly known. Historically, only one species, “Hippotherium” antelopinum Falconer & Cautley, 1849, was described from the Upper Siwaliks. Here, we present the frst evidence of Eurygnathohippus Van Hoepen, 1930, a lineage hitherto only known from Africa, in the Upper Siwaliks during the late Pliocene. Morphologically, the South Asian Eurygnathohippus is most similar to Eurygnathohippus hasumense (Eisenmann, 1983) from Afar, Ethiopia, a species with a similar temporal range. -

To My Mind, Not Due So Much to the Lack of Change in the Teeth As to The
19291 Matthew, Critical Observations upon Siwalik Mammals 553 to my mind, not due so much to the lack of change in the teeth as to the rapidity of change in the skull, and the whole family derives from late Miocene paleomerycines, an antiquity decidedly less than most mam- malian families. According to Pilgrim' the Giraffidae afford " one of our most complete proofs for the Pontian age of the Dhok Pathan." But his argument will not stand analysis. "There we find side by side with G. attica is in fact distinctly smaller and Giraffa punjabiensis, paralleled by more brachydont than punjabiensis. Giraffa attica at Pikermi, the large genus What Pilgrim called 'Helladotherium' HeUadotherium allied to the Pikermi grande is in fact Lydekker's Hydaspi- species H. duvernoyi and the almost therium 7negacephalum. HeUadotherium equally large Vishnutherium. duvernoyi is much more primitive; com- pares with "Vishnutherium," but then " Vishnutherium" occurs in both the middle and lower Siwalik. "The palaeotragine group, including the Giraffokeryx is a member of it, and is genera Palzeotragus, Sanwtherium and considered by Bohlin to be a species of Alcicephalus, has not so far been Paleotragus. One species of Giraffokeryx, recognized in India. however, if not both, carries four horns, and is generically distinct from Paleo- tragus, although clearly related. "On the other hand, allied forms posses- As we should expect if the Dhok Pathan sing a complex horn development occur and Perim Island beds are later than in the genus Hyda pitherium in the Dhok Pikermi, Samos or Northern China. Pathan zone, and as Bramatherium in the slightly older Perim Island beds. -

The Phylogeny and Taxonomy of Hippopotamidae (Mammalia: Artiodactyla): a Review Based on Morphology and Cladistic Analysis
Blackwell Science, LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082The Lin- nean Society of London, 2005? 2005 143? 126 Original Article J.-R. BOISSERIEHIPPOPOTAMIDAE PHYLOGENY AND TAXONOMY Zoological Journal of the Linnean Society, 2005, 143, 1–26. With 11 figures The phylogeny and taxonomy of Hippopotamidae (Mammalia: Artiodactyla): a review based on morphology and cladistic analysis JEAN-RENAUD BOISSERIE1,2* 1Laboratoire de Géobiologie, Biochronologie et Paléontologie Humaine, UMR CNRS 6046, Université de Poitiers, 40 avenue du Recteur, Pineau 86022 Cedex, France 2Laboratory for Human Evolutionary Studies, Department of Integrative Biology, Museum of Vertebrate Zoology, University of California at Berkeley, 3101 Valley Life Science Building, Berkeley, CA 94720-3160, USA Received August 2003; accepted for publication June 2004 The phylogeny and taxonomy of the whole family Hippopotamidae is in need of reconsideration, the present confu- sion obstructing palaeoecology and palaeobiogeography studies of these Neogene mammals. The revision of the Hip- popotamidae initiated here deals with the last 8 Myr of African and Asian species. The first thorough cladistic analysis of the family is presented here. The outcome of this analysis, including 37 morphological characters coded for 15 extant and fossil taxa, as well as non-coded features of mandibular morphology, was used to reconstruct broad outlines of hippo phylogeny. Distinct lineages within the paraphyletic genus Hexaprotodon are recognized and char- acterized. In order to harmonize taxonomy and phylogeny, two new genera are created. The genus name Choeropsis is re-validated for the extant Liberian hippo. The nomen Hexaprotodon is restricted to the fossil lineage mostly known in Asia, but also including at least one African species. -

A 3D Stereotactic Atlas of the Adult Human Skull Base Wieslaw L
Nowinski and Thaung Brain Inf. (2018) 5:1 https://doi.org/10.1186/s40708-018-0082-1 Brain Informatics ORIGINAL RESEARCH Open Access A 3D stereotactic atlas of the adult human skull base Wieslaw L. Nowinski1,2* and Thant S. L. Thaung3 Abstract Background: The skull base region is anatomically complex and poses surgical challenges. Although many textbooks describe this region illustrated well with drawings, scans and photographs, a complete, 3D, electronic, interactive, real- istic, fully segmented and labeled, and stereotactic atlas of the skull base has not yet been built. Our goal is to create a 3D electronic atlas of the adult human skull base along with interactive tools for structure manipulation, exploration, and quantifcation. Methods: Multiple in vivo 3/7 T MRI and high-resolution CT scans of the same normal, male head specimen have been acquired. From the scans, by employing dedicated tools and modeling techniques, 3D digital virtual models of the skull, brain, cranial nerves, intra- and extracranial vasculature have earlier been constructed. Integrating these models and developing a browser with dedicated interaction, the skull base atlas has been built. Results: This is the frst, to our best knowledge, truly 3D atlas of the adult human skull base that has been created, which includes a fully parcellated and labeled brain, skull, cranial nerves, and intra- and extracranial vasculature. Conclusion: This atlas is a useful aid in understanding and teaching spatial relationships of the skull base anatomy, a helpful tool to generate teaching materials, and a component of any skull base surgical simulator. Keywords: Skull base, Electronic atlas, Digital models, Skull, Brain, Stereotactic atlas 1 Introduction carotid arteries, among others. -

Faunal and Environmental Change in the Late Miocene Siwaliks of Northern Pakistan
Copyright ( 2002, The Paleontological Society Faunal and environmental change in the late Miocene Siwaliks of northern Pakistan John C. Barry, MicheÁle E. Morgan, Lawrence J. Flynn, David Pilbeam, Anna K. Behrensmeyer, S. Mahmood Raza, Imran A. Khan, Catherine Badgley, Jason Hicks, and Jay Kelley Abstract.ÐThe Siwalik formations of northern Pakistan consist of deposits of ancient rivers that existed throughout the early Miocene through the late Pliocene. The formations are highly fossil- iferous with a diverse array of terrestrial and freshwater vertebrates, which in combination with exceptional lateral exposure and good chronostratigraphic control allows a more detailed and tem- porally resolved study of the sediments and faunas than is typical in terrestrial deposits. Conse- quently the Siwaliks provide an opportunity to document temporal differences in species richness, turnover, and ecological structure in a terrestrial setting, and to investigate how such differences are related to changes in the ¯uvial system, vegetation, and climate. Here we focus on the interval between 10.7 and 5.7 Ma, a time of signi®cant local tectonic and global climatic change. It is also the interval with the best temporal calibration of Siwalik faunas and most comprehensive data on species occurrences. A methodological focus of this paper is on controlling sampling biases that confound biological and ecological signals. Such biases include uneven sampling through time, differential preservation of larger animals and more durable skeletal elements, errors in age-dating imposed by uncertainties in correlation and paleomagnetic timescale calibrations, and uneven tax- onomic treatment across groups. We attempt to control for them primarily by using a relative-abun- dance model to estimate limits for the ®rst and last appearances from the occurrence data. -

Extraoral Anatomy in CBCT - Michael M
126 RESEARCH AND SCIENCE Thomas von Arx1 Scott Lozanoff2 Extraoral anatomy in CBCT - Michael M. Bornstein3,4 a literature review 1 Department of Oral Surgery and Stomatology, School of Dental Medicine, University of Bern, Switzerland Part 2: Zygomatico-orbital region 2 Department of Anatomy, Biochemistry and Physiology, John A. Burns School of Medi- cine, University of Hawaii, Honolulu, USA 3 Oral and Maxillofacial Radiol- ogy, Applied Oral Sciences KEYWORDS and Community Dental Care, Anatomy Faculty of Dentistry, The Uni- CBCT versity of Hong Kong, Prince Zygomatic bone Philip Dental Hospital, Hong Orbital cavity Kong SAR, China 4 Department of Oral Health & Medicine, University Center for Dental Medicine Basel UZB, University of Basel, Basel, Switzerland SUMMARY CORRESPONDENCE Prof. Dr. Thomas von Arx This second article about extraoral anatomy as noid bone along the lateral orbital wall. Each of Klinik für Oralchirurgie und seen in cone beam computed tomography (CBCT) the three surfaces of the zygomatic bone displays Stomatologie images presents a literature review of the zygo- foramina that transmit neurovascular structures. Zahnmedizinische Kliniken matico-orbital region. The latter bounds the The orbital cavity is located immediately above der Universität Bern maxillary sinus superiorly and laterally. Since the maxillary sinus from which it is separated Freiburgstrasse 7 CH-3010 Bern pathologic changes of the maxillary sinus are a only by a thin bony plate simultaneously serving Tel. +41 31 632 25 66 frequent indication for three-dimensional radi- as the orbital floor and the roof of the maxillary Fax +41 31 632 25 03 ography, the contiguous orbital cavity and the sinus. -

Harrison CV June 2021
June 1, 2021 Terry Harrison CURRICULUM VITAE CONTACT INFORMATION * Center for the Study of Human Origins Department of Anthropology 25 Waverly Place New York University New York, NY 10003-6790, USA 8 [email protected] ) 212-998-8581 WEB LINKS http://as.nyu.edu/faculty/terry-harrison.html https://wp.nyu.edu/csho/people/faculty/terry_harrison/ https://nyu.academia.edu/TerryHarrison http://orcid.org/0000-0003-4224-0152 zoobank.org:author:43DA2256-CF4D-476F-8EA8-FBCE96317505 ACADEMIC BACKGROUND Graduate: 1978–1982: Doctor of Philosophy. Department of Anthropology, University College London, London. Doctoral dissertation: Small-bodied Apes from the Miocene of East Africa. 1981–1982: Postgraduate Certificate of Education. Institute of Education, London University, London. Awarded with Distinction. Undergraduate: 1975–1978: Bachelor of Science. Department of Anthropology, University College London, London. First Class Honours. POSITIONS 2014- Silver Professor, Department of Anthropology, New York University. 2003- Director, Center for the Study of Human Origins, New York University. 1995- Professor, Department of Anthropology, New York University. 2010-2016 Chair, Department of Anthropology, New York University. 1995-2010 Associate Chair, Department of Anthropology, New York University. 1990-1995 Associate Professor, Department of Anthropology, New York University. 1984-1990 Assistant Professor, Department of Anthropology, New York University. HONORS & AWARDS 1977 Rosa Morison Memorial Medal and Prize, University College London. 1978 Daryll Forde Award, University College London. 1989 Golden Dozen Award for excellence in teaching, New York University. 1996 Golden Dozen Award for excellence in teaching, New York University. 2002 Distinguished Teacher Award, New York University. 2006 Fellow, American Association for the Advancement of Science. -
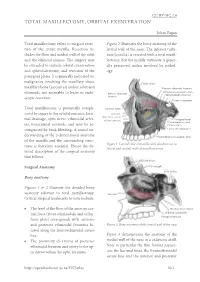
Total Maxillectomy and Orbital Exenteration
CC-BY-NC 3.0 TOTAL MAXILLECTOMY, ORBITAL EXENTERATION Johan Fagan Total maxillectomy refers to surgical resec- Figure 2 illustrates the bony anatomy of the tion of the entire maxilla. Resection in- lateral wall of the nose. The inferior turbi- cludes the floor and medial wall of the orbit nate (concha) is resected with a total maxil- and the ethmoid sinuses. The surgery may lectomy, but the middle turbinate is gener- be extended to include orbital exenteration ally preserved, unless involved by pathol- and sphenoidectomy, and resection of the ogy. pterygoid plates. It is generally indicated for malignancy involving the maxillary sinus, Frontal sinus maxillary bone (sarcomas) and/or orbit and Posterior ethmoidal foramen Orbital process palatine bone ethmoids, not amenable to lesser or endo- Anterior ethmoidal Sphenopalatine foramen scopic resection. foramen Foramen rotundum Total maxillectomy is potentially compli- Lacrimal fossa cated by injury to the orbital contents, lacri- Uncinate Max sinus ostium mal drainage, optic nerve, ethmoidal arter- Inferior turbinate Pterygoid canal Pterygopalatine canal ies, intracranial contents, and may be ac- Palatine bone companied by brisk bleeding. A sound un- Lateral pterygoid plate derstanding of the 3-dimensional anatomy Pyramidal process palatine bone of the maxilla and the surrounding struc- Figure 1: Lateral view of maxilla with windows cut in tures is therefore essential. Hence the de- lateral and medial walls of maxillary sinus tailed description of the surgical anatomy that follows. Frontal