Adapting to Climate Change at Olympic National Forest And
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Socioeconomic Monitoring of the Olympic National Forest and Three Local Communities
NORTHWEST FOREST PLAN THE FIRST 10 YEARS (1994–2003) Socioeconomic Monitoring of the Olympic National Forest and Three Local Communities Lita P. Buttolph, William Kay, Susan Charnley, Cassandra Moseley, and Ellen M. Donoghue General Technical Report United States Forest Pacific Northwest PNW-GTR-679 Department of Service Research Station July 2006 Agriculture The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the National Forests and National Grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable, sex, marital status, familial status, parental status, religion, sexual orientation, genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all pro- grams.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD). To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, 1400 Independence Avenue, SW, Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. -

Snowmobiles in the Wilderness
Snowmobiles in the Wilderness: You can help W a s h i n g t o n S t a t e P a r k s A necessary prohibition Join us in safeguarding winter recreation: Each year, more and more people are riding snowmobiles • When riding in a new area, obtain a map. into designated Wilderness areas, which is a concern for • Familiarize yourself with Wilderness land managers, the public and many snowmobile groups. boundaries, and don’t cross them. This may be happening for a variety of reasons: many • Carry the message to clubs, groups and friends. snowmobilers may not know where the Wilderness boundaries are or may not realize the area is closed. For more information about snowmobiling opportunities or Wilderness areas, please contact: Wilderness…a special place Washington State Parks and Recreation Commission (360) 902-8500 Established by Congress through the Wilderness Washington State Snowmobile Association (800) 784-9772 Act of 1964, “Wilderness” is a special land designation North Cascades National Park (360) 854-7245 within national forests and certain other federal lands. Colville National Forest (509) 684-7000 These areas were designated so that an untouched Gifford Pinchot National Forest (360) 891-5000 area of our wild lands could be maintained in a natural Mt. Baker-Snoqualmie National Forest (425) 783-6000 state. Also, they were set aside as places where people Mt. Rainier National Park (877) 270-7155 could get away from the sights and sounds of modern Okanogan-Wenatchee National Forest (509) 664-9200 civilization and where elements of our cultural history Olympic National Forest (360) 956-2402 could be preserved. -

Land Areas of the National Forest System, As of September 30, 2019
United States Department of Agriculture Land Areas of the National Forest System As of September 30, 2019 Forest Service WO Lands FS-383 November 2019 Metric Equivalents When you know: Multiply by: To fnd: Inches (in) 2.54 Centimeters Feet (ft) 0.305 Meters Miles (mi) 1.609 Kilometers Acres (ac) 0.405 Hectares Square feet (ft2) 0.0929 Square meters Yards (yd) 0.914 Meters Square miles (mi2) 2.59 Square kilometers Pounds (lb) 0.454 Kilograms United States Department of Agriculture Forest Service Land Areas of the WO, Lands National Forest FS-383 System November 2019 As of September 30, 2019 Published by: USDA Forest Service 1400 Independence Ave., SW Washington, DC 20250-0003 Website: https://www.fs.fed.us/land/staff/lar-index.shtml Cover Photo: Mt. Hood, Mt. Hood National Forest, Oregon Courtesy of: Susan Ruzicka USDA Forest Service WO Lands and Realty Management Statistics are current as of: 10/17/2019 The National Forest System (NFS) is comprised of: 154 National Forests 58 Purchase Units 20 National Grasslands 7 Land Utilization Projects 17 Research and Experimental Areas 28 Other Areas NFS lands are found in 43 States as well as Puerto Rico and the Virgin Islands. TOTAL NFS ACRES = 192,994,068 NFS lands are organized into: 9 Forest Service Regions 112 Administrative Forest or Forest-level units 503 Ranger District or District-level units The Forest Service administers 149 Wild and Scenic Rivers in 23 States and 456 National Wilderness Areas in 39 States. The Forest Service also administers several other types of nationally designated -

Skagit River Emergency Bank Stabilization And
United States Department of the Interior FISH AND WILDLIFE SERVICE Washington Fish and Wildlife Office 510 Desmond Dr. SE, Suite 102 Lacey, Washington 98503 In Reply Refer To: AUG 1 6 2011 13410-2011-F-0141 13410-2010-F-0175 Daniel M. Mathis, Division Adnlinistrator Federal Highway Administration ATIN: Jeff Horton Evergreen Plaza Building 711 Capitol Way South, Suite 501 Olympia, Washington 98501-1284 Michelle Walker, Chief, Regulatory Branch Seattle District, Corps of Engineers ATIN: Rebecca McAndrew P.O. Box 3755 Seattle, Washington 98124-3755 Dear Mr. Mathis and Ms. Walker: This document transmits the U.S. Fish and Wildlife Service's Biological Opinion (Opinion) based on our review of the proposed State Route 20, Skagit River Emergency Bank Stabilization and Chronic Environmental Deficiency Project in Skagit County, Washington, and its effects on the bull trout (Salve linus conjluentus), marbled murrelet (Brachyramphus marmoratus), northern spotted owl (Strix occidentalis caurina), and designated bull trout critical habitat, in accordance with section 7 of the Endangered Species Act of 1973, as amended (16 U.S.C. 1531 et seq.) (Act). Your requests for initiation of formal consultation were received on February 3, 2011, and January 19,2010. The Federal Highway Administration (FHWA) and U.S. Army Corps of Engineers - Seattle District (Corps) provided information in support of "may affect, likely to adversely affect" determinations for the bull trout and designated bull trout critical habitat. On April 13 and 14, 2011, the FHWA and Corps gave their consent for addressing potential adverse effects to the bull trout and bull trout critical habitat with a single Opinion. -

Chehalis River
Northwest Area Committee OCTOBER 2015 CHEHALIS RIVER Geographic Response Plan (CHER GRP) 1 This page was intentionally left blank. 2 CHEHALIS RIVER GRP OCTOBER 2015 CHEHALIS RIVER Geographic Response Plan (CHER GRP) October 2015 3 CHEHALIS RIVER GRP OCTOBER 2015 Spill Response Contact Sheet Required Notifications for Oil Spills and Hazardous Substance Releases Federal Notification - National Response Center (800) 424-8802* State Notification - Washington Emergency Management (800) 258-5990* Division - Other Contact Numbers - U.S. Environmental Protection Agency Washington State Region 10 - Spill Response (206) 553-1263* Dept Archaeology & Hist Preserv (360) 586-3065 - Washington Ops Office (360) 753-9437 Dept of Ecology - Oregon Ops Office (503) 326-3250 - Headquarters (Lacey) (360) 407-6000 - RCRA/CERCLA Hotline (800) 424-9346 - SW Regional Office (Lacey) (360) 407-6300 - Public Affairs (206) 553-1203 Dept of Fish and Wildlife (360) 902-2200 - Emergency HPA Assistance (360) 902-2537* U.S. Coast Guard -Oil Spill Team (360) 534-8233* Sector Columbia River Dept of Health (Drinking Water) (800) 521-0323 - Emergency / Watchstander (503) 861-2242* - After normal business hours (877) 481-4901 - Command Center (503) 861-6211* Dept of Natural Resources (360) 902-1064 - Incident Management Division (503) 861-6477 - After normal business hours (360) 556-3921 - Station Grays Harbor (360) 268-0121* Dept of Transportation (360) 705-7000 13th Coast Guard District (800) 982-8813 State Parks & Rec Commission (360) 902-8613 National Strike Force (252) 331-6000 State Patrol - District 1 (253) 538-3240 Coordination - Pacific Strike Center Team (415) 883-3311 State Patrol - District 5 (360) 449-7909 State Patrol - District 8 (360) 473-0172 National Oceanic Atmospheric Administration Scientific Support Coordinator (206) 526-6829 Tribal Contacts Weather (206) 526-6087 Chehalis Confederated Tribes (360) 273-5911 - Cultural Resources Ext. -

Elma Washington ~ Now and Then
Area Information Elma, WA around 1898 to 1900 Main Street Looking East From 5th Street By Luke Bailey Image 2010 “Satsop Development Park was the winner of the Governor’s 2004 Economic Development Award for Infrastructure.” Innovative features in a unique setting Find out more today about Satsop Development Park, recently designated one of the top 10 industrial and office parks in Washington state for 2005 by Washington CEO magazine. Exceptional features include: Diversely routed fiber optics Only 5 minutes from Elma off Highway 12 Unlimited bandwidth potential Intercom phone system serving 20,000 ports 24/7 network monitoring, resolution & assistance Exceptional security in location and buildings Uninterruptible AC power system Room for all types of businesses to grow Back-up generators Regional workforce of over 68,000 people Visit our website: www.satsop.com 1.866.5SATSOP or 360.482.1600 [email protected] Grays Harbor Public Development Authority An economic powerhouse creating jobs and investment in Grays Harbor TABLE OF CONTENTS Welcome From the Elma Chamber of Commerce ......4 Elma Government & Community Services ........... 5 The Elma Microtel Inn & Suites .......................... 6 Chamber Parks & Recreation Elma & East County ....... 7-13 Of Elma Washington ~ Now and Then ............. 15-22 ELMA SCHOOL DISTRICT #68 ...................... 22-23 Commerce Quarter Midget Racing ......................... 25 Welcomes Sprint Car Racing and the World of Outlaws .... 26 You to Community Events ........................... 27-28 The Elma Murals of Elma ............................. 30-31 Map of Elma ................................... 32 Community Census and Demographics ~ Elma ............. 33-35 Business & Shopping Directory .............. 37-43 The Elma Chamber of Commerce Expresses Special Tom Rose Thanks to All Those Who Helped Make This 1912 Photo of Elma from atop Strawberry Publication Possible. -

Coastal Recovery Unit Implementation Plan for Bull Trout (Salvelinus Confluentus)
U.S. Fish & Wildlife Service Coastal Recovery Unit Implementation Plan for Bull Trout (Salvelinus confluentus) Top left: Clackamas bull trout reintroduction, Clackamas River, Oregon. David Herasimtschuk, Freshwaters Illustrated; Top, right: Glines Canyon Dam removal, Elwha River, Washington. John Gussman, Doubleclick Productions; Center: South Fork Skagit River and Skagit Bay, Washington. City of Seattle; Bottom: Riverscape surveys, East Fork Quinault River, Washington. National Park Service, Olympic National Park Coastal Recovery Unit Implementation Plan for Bull Trout (Salvelinus confluentus) September 2015 Prepared by U.S. Fish and Wildlife Service Washington Fish and Wildlife Office Lacey, Washington and Oregon Fish and Wildlife Office Portland, Oregon Table of Contents Introduction ................................................................................................................................. A-1 Current Status of Bull Trout in the Coastal Recovery Unit ........................................................ A-6 Factors Affecting Bull Trout in the Coastal Recovery Unit ....................................................... A-8 Ongoing Coastal Recovery Unit Conservation Measures (Summary) ..................................... A-33 Research, Monitoring, and Evaluation ..................................................................................... A-38 Recovery Measures Narrative ................................................................................................... A-39 Implementation Schedule for -
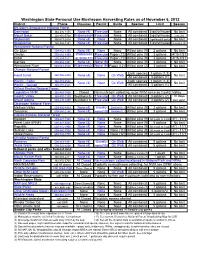
Washington State Personal Use Mushroom Harvesting Rules As of November 6, 2012 District Phone Closures Permit Guide Species Limit Season Mt
Washington State Personal Use Mushroom Harvesting Rules as of November 6, 2012 District Phone Closures Permit Guide Species Limit Season Mt. Baker-Snoqualmie National Forest Darrington 360-436-1155 None (4) Free use None All combined 5 gal/yr/house No limit Mount Baker 360-856-5700 Wilderness(4) Free use None All combined 5 gal/yr/house 2 weeks Skykomish 360-677-2414 None (4) None None All combined 1 gal/day&5 gal/yr No limit Snoqualmie 425-888-1421 None (4) Free use None All combined 5 gal/yr/house 2 weeks (8) Wenatchee National Forest Cle Elum 509-852-1100 None (4) None None All but pine (9) 2 gallons No limit Chelan 509-682-4900 Wilderness Free use Paper (12) All but pine (9) 3 gallons 4/15-7/31 Entiat 509-784-1511 &LSR(4,11) Free use Paper (12) All but pine (9) 3 gallons 4/15-7/31 Naches 509-653-1401 None (4) None (9) None All but pine (9) 3 gallons No limit Wenatchee River 509-548-2550 Wilderness(4) None (9) Paper (12) All but pine (2) 3 gallons No limit Olympic National Forest Each species 1 gallon (1,3) Hood Canal 360-765-2200 None (4) None On Web No limit All combined 3 gallons (1) Pacific - Forks 360-374-6522 Each species 1 gallon (1,3) None (4) None On Web No limit Pacific - Quinalt 360-288-2525 All combined 3 gallons (1) Gifford Pinchot National Forest Legislative NVM 360-449-7800 Closed No mushroom collecting, outer NVM same as Cowlitz Valley Cowlitz Valley 360-497-1100 SeeMap(4,5) Free use On Web All combined 3 gallons (2) 10 days Mount Adams 509-395-3400 SeeMap(4,5) Free use On Web All combined 3 gallons (2) per year Okanogan -

Public Law 94-527 94Th Congress an Act to Amend the National Trails System Act (82 Stat
PUBLIC LAW 94-527—OCT. 17, 1976 90 STAT. 2481 Public Law 94-527 94th Congress An Act To amend the National Trails System Act (82 Stat. 919), and for other purposes. Oct. 17, 1976 [S. 2112] Be it enacted hy the Senate and House of Representatives of the United States of America in Congress assembled, That the National National scenic Trails System Act (82 Stat. 919; 16 U.S.C. 1241 et seq.) is amended trails, as follows: In section 5(c), add the following new paragraphs; 16 USC 1244. "(15) Bartram Trail, extending through the States of Georgia, North Carolina, South Carolina, Alabama, Florida, Louisiana, Mis sissippi, and Tennessee. "(16) Daniel Boone Trail, extending from the vicinity of States- ville. North Carolina, to Fort Boonesborough State Park, Kentucky. "(17) Desert Trail, extending from the Canadian border through parts of Idaho, Washington, Oregon, Nevada, California, and Ari zona, to the Mexican border. "(18) Dominguez-Escalante Trail, extending approximately two thousand miles along the route of the 1776 expedition led by Father Francisco Atanasio Dominguez and Father Silvestre Velez de Escalante, originating in Santa Fe, New Mexico; proceeding north west along the San Juan, Dolores, Gunnison, and White Rivers in Colorado; thence westerly to Utah Lake; thence southward to Arizona and returning to Santa Fe. "(19) Florida Trail, extending north from Everglades National Park, including the Big Cypress Swamp, the Kissimme Prairie, the Withlacoochee State Forest, Ocala National Forest, Osceola National Forest, and Black Water River State Forest, said completed trail to be approximately one thousand three hundred miles long, of which over four hundred miles of trail have already been built. -
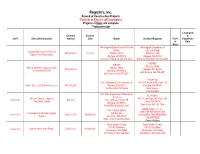
Projects *Projects in Red Are Still in Progress Projects in Black Are Complete **Subcontractor
Rognlin’s, Inc. Record of Construction Projects *Projects in Red are still in progress Projects in black are complete **Subcontractor % Complete Contract Contract & Job # Description/Location Amount Date Owner Architect/Engineer Class Completion of Date Work Washington Department of Fish and Washington Department of Wildlife Fish and Wildlife Log Jam Materials for East Fork $799,000.00 01/11/21 PO Box 43135 PO Box 43135 Satsop River Restoration Olympia, WA 98504 Olympia, WA 98504 Adrienne Stillerman 360.902.2617 Adrienne Stillerman 360.902.2617 WSDOT WSDOT PO Box 47360 SR 8 & SR 507 Thurston County PO Box 47360 $799,000.00 Olympia, WA 98504 Stormwater Retrofit Olympia, WA 98504 John Romero 360.570.6571 John Romero 360.570.6571 Parametrix City of Olympia 601 4th Avennue E. 1019 39th Avenue SE, Suite 100 Water Street Lift Station Generator $353,952.76 Olympia, WA 98501 Puyallup, WA 98374 Jim Rioux 360-507-6566 Kevin House 253.604.6600 WA State Department of Enterprise SCJ Alliance Services 14th Ave Tunnel – Improve 8730 Tallon Lane NE, Suite 200 20-80-167 $85,000 1500 Jefferson Street SE Pedestrian Safety Lacey, WA 98516 Olympia, WA 98501 Ross Jarvis 360-352-1465 Bob Willyerd 360.407.8497 ABAM Engineers, Inc. Port of Grays Harbor 33301 9th Ave S Suite 300 Terminals 3 & 4 Fender System PO Box 660 20-10-143 $395,118.79 12/08/2020 Federal Way, WA 98003-2600 Repair Aberdeen, WA 98520 (206) 357-5600 Mike Johnson 360.533.9528 Robert Wallace Grays Harbor County Grays Harbor County 100 W. -

Land and Resource Management Plan
United States Department of Land and Resource Agriculture Forest Service Management Plan Pacific Northwest Region 1990 Olympic National Forest I,,; ;\'0:/' "\l . -'. \.. \:~JK~~'.,;"> .. ,. :~i;/i- t~:.(~#;~.. ,':!.\ ," "'~.' , .~, " ,.. LAND AND RESOURCE MANAGEMENT PLAN for the OLYMPIC NATIONAL FOREST PACIFIC NORTHWEST REGION PREFACE Preparation of a Land and Resource Management Plan (Forest Plan) for the Olympic National Forest is required by the Forest and Rangeland Renewable Resources Planning Act (RPA) as amended by the National Forest Management Act (NFMA). Regulations developed under the RPA establish a process for developing, adopting, and revising land and resource Plans for the National Forest System (36 CFR 219). The Plan has also been developed in accordance with regulations (40 CFR 1500) for implementing the National Environmental Policy Act of 1969 (NEPA). Because this Plan is considered a major Federal action significantly affecting the quality of the human environment, a detailed statement (environmental impact statement) has been prepared as required by NEPA. The Forest Plan represents the implementation of the Preferred Alternative as identified in the Final Environmental Impact Statement (FEIS) for the Forest Plan. If any particular provision of this Forest Plan, or application of the action to any person or circumstances is found to be invalid, the remainder of this Forest Plan and the application of that provision to other persons or circumstances shall not be affected. Information concerning this plan can be obtained -

Recreation • NORTH Knowledge Our of Best the to Accurate Is Herein Presented Information All
December 2008 December not responsible for any inaccuracies of map or hardship/injury due to same. same. to due hardship/injury or map of inaccuracies any for responsible not please check for current conditions before planning your trip. Map authors authors Map trip. your planning before conditions current for check please and sources as of printing date. Road conditions change periodically, so so periodically, change conditions Road date. printing of as sources and Mason County Recreation • NORTH knowledge our of best the to accurate is herein presented information All Mason County is home to a variety of outdoor recreation, thanks to our towering mountains, huge tracts of national forest, and hundreds Mason County Transit is Bicycle Friendly! of miles of salt and freshwater shoreline. A variety of bicycle routes are LEGEND highlighted with convenient map keys. Additionally, we’ve identified If roads get too busy or you grow too tired, remember that all Mason simply a matter of judgement regarding safety and passenger loads. BIKEWAYS many scenic and popular launches for kayaking, boating and fishing. For POINTS OF INTEREST County Transit buses provide bike racks. Rarely, but sometimes, you may be asked to take the next bus. If you bring your bicycle on board, please be responsible for its handling those who want to escape further into the forest, this map will show you Bike Routes Convenient, secure and easy-to-use, bicycle racks become a bicyclist’s and security in the wheelchair securement area throughout the trip. some of our favorite trails and trailheads for hiking and exploring. State Public Restroom Visitor Information Heavy Trac ? best friend when weather conditions worsen or visibility lessens due and county parks for camping or outdoor play, Mason Transit bicycle- Schedule Information • 360-426-5033 Boat Launch to darkness, mist or fog.