Genomewide View of Gene Silencing by Small Interfering Rnas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Chapter 14: Functional Genomics Learning Objectives
Chapter 14: Functional Genomics Learning objectives Upon reading this chapter, you should be able to: ■ define functional genomics; ■ describe the key features of eight model organisms; ■ explain techniques of forward and reverse genetics; ■ discuss the relation between the central dogma and functional genomics; and ■ describe proteomics-based approaches to functional genomics. Outline : Functional genomics Introduction Relation between genotype and phenotype Eight model organisms E. coli; yeast; Arabidopsis; C. elegans; Drosophila; zebrafish; mouse; human Functional genomics using reverse and forward genetics Reverse genetics: mouse knockouts; yeast; gene trapping; insertional mutatgenesis; gene silencing Forward genetics: chemical mutagenesis Functional genomics and the central dogma Approaches to function; Functional genomics and DNA; …and RNA; …and protein Proteomic approaches to functional genomics CASP; protein-protein interactions; protein networks Perspective Albert Blakeslee (1874–1954) studied the effect of altered chromosome numbers on the phenotype of the jimson-weed Datura stramonium, a flowering plant. Introduction: Functional genomics Functional genomics is the genome-wide study of the function of DNA (including both genes and non-genic regions), as well as RNA and proteins encoded by DNA. The term “functional genomics” may apply to • the genome, transcriptome, or proteome • the use of high-throughput screens • the perturbation of gene function • the complex relationship of genotype and phenotype Functional genomics approaches to high throughput analyses Relationship between genotype and phenotype The genotype of an individual consists of the DNA that comprises the organism. The phenotype is the outward manifestation in terms of properties such as size, shape, movement, and physiology. We can consider the phenotype of a cell (e.g., a precursor cell may develop into a brain cell or liver cell) or the phenotype of an organism (e.g., a person may have a disease phenotype such as sickle‐cell anemia). -

Split Deoxyribozyme Probe for Efficient Detection of Highly Structured RNA Targets
University of Central Florida STARS Honors Undergraduate Theses UCF Theses and Dissertations 2018 Split Deoxyribozyme Probe For Efficient Detection of Highly Structured RNA Targets Sheila Raquel Solarez University of Central Florida Part of the Biochemistry Commons, and the Biology Commons Find similar works at: https://stars.library.ucf.edu/honorstheses University of Central Florida Libraries http://library.ucf.edu This Open Access is brought to you for free and open access by the UCF Theses and Dissertations at STARS. It has been accepted for inclusion in Honors Undergraduate Theses by an authorized administrator of STARS. For more information, please contact [email protected]. Recommended Citation Solarez, Sheila Raquel, "Split Deoxyribozyme Probe For Efficient Detection of Highly Structured RNA Targets" (2018). Honors Undergraduate Theses. 311. https://stars.library.ucf.edu/honorstheses/311 SPLIT DEOXYRIBOZYME PROBE FOR EFFICIENT DETECTION OF HIGHLY STRUCTURED RNA TARGETS By SHEILA SOLAREZ A thesis submitted in partial fulfillment of the requirements for the Honors in the Major Program in Biological Sciences in the College of Sciences and the Burnett Honors College at the University of Central Florida Orlando, Florida Spring Term, 2018 Thesis Chair: Yulia Gerasimova, PhD ABSTRACT Transfer RNAs (tRNAs) are known for their role as adaptors during translation of the genetic information and as regulators for gene expression; uncharged tRNAs regulate global gene expression in response to changes in amino acid pools in the cell. Aminoacylated tRNAs play a role in non-ribosomal peptide bond formation, post-translational protein labeling, modification of phospholipids in the cell membrane, and antibiotic biosynthesis. [1] tRNAs have a highly stable structure that can present a challenge for their detection using conventional techniques. -

Mrna Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection
International Journal of Molecular Sciences Review mRNA Vaccine Era—Mechanisms, Drug Platform and Clinical Prospection 1, 1, 2 1,3, Shuqin Xu y, Kunpeng Yang y, Rose Li and Lu Zhang * 1 State Key Laboratory of Genetic Engineering, Institute of Genetics, School of Life Science, Fudan University, Shanghai 200438, China; [email protected] (S.X.); [email protected] (K.Y.) 2 M.B.B.S., School of Basic Medical Sciences, Peking University Health Science Center, Beijing 100191, China; [email protected] 3 Shanghai Engineering Research Center of Industrial Microorganisms, Shanghai 200438, China * Correspondence: [email protected]; Tel.: +86-13524278762 These authors contributed equally to this work. y Received: 30 July 2020; Accepted: 30 August 2020; Published: 9 September 2020 Abstract: Messenger ribonucleic acid (mRNA)-based drugs, notably mRNA vaccines, have been widely proven as a promising treatment strategy in immune therapeutics. The extraordinary advantages associated with mRNA vaccines, including their high efficacy, a relatively low severity of side effects, and low attainment costs, have enabled them to become prevalent in pre-clinical and clinical trials against various infectious diseases and cancers. Recent technological advancements have alleviated some issues that hinder mRNA vaccine development, such as low efficiency that exist in both gene translation and in vivo deliveries. mRNA immunogenicity can also be greatly adjusted as a result of upgraded technologies. In this review, we have summarized details regarding the optimization of mRNA vaccines, and the underlying biological mechanisms of this form of vaccines. Applications of mRNA vaccines in some infectious diseases and cancers are introduced. It also includes our prospections for mRNA vaccine applications in diseases caused by bacterial pathogens, such as tuberculosis. -

Patenting Interfering RNA
Patenting Interfering RNA J. Douglas Schultz SPE Art Unit 1635 (571) 272-0763 [email protected] Oligonucleotide Inhibitors: Mechanisms of Action RNAi - Mechanism of Action • dsRNA induces sequence-specific degradation of homologous gene transcripts • RISC metabolizes dsRNA to small 21-23- nucleotide siRNAs – RISC contains dsRNase (“Dicer”), ssRNase (Argonaut 2 or Ago2) • RISC utilizes antisense strand as “guide” to find cleavable target siRNA Mechanism of Action miRNA Mechanism of Action Interfering RNA Glossary of Terms • RNAi – RNA interference • dsRNA – double stranded RNA • siRNA – small interfering RNA, double stranded, 21-23 nucleotides • shRNA – short hairpin RNA (doubled stranded by virtue of a ssRNA folding back on itself) • miRNA – micro RNA • RISC – RNA-induced silencing complex – Dicer – RNase endonuclease siRNA miRNA • Exogenously delivered • Endogenously produced • 21-23mer dsRNA • 21-23mer dsRNA • Acts through RISC • Acts through RISC • Induces homologous target • Induces homologous target cleavage cleavage • Perfect sequence match • Imperfect sequence match – Results in target degradation – Results in translation arrest RNAi Patentability issues Sample Claims: • A siRNA that inhibits expression of a nucleic acid encoding protein X. OR • A siRNA comprising a 2’-modification, wherein said modification comprises 2’-fluoro, 2’-O-methyl, or 2’- deoxy. (Note: no target recited) OR • A method of reducing tumor cell growth comprising administering siRNA targeting protein X. RNAi Patentability Issues 35 U.S.C. 101 – Utility • Credible/Specific/Substantial/Well Established. • Used to attempt modulation of gene expression in human diseases • Routinely investigate gene function in a high throughput fashion or to (see Rana RT, Nat. Rev. Mol. Cell Biol. 2007, Vol. 8:23-36). -

Andrey L. Karamyshev
CURRICULUM VITAE Andrey L. Karamyshev Assistant Professor Department of Cell Biology and Biochemistry! Texas Tech University Health Sciences Center! 3601 4th Street, Mail Stop 6540! Lubbock, TX 79430 Phone: (806) 743-4102 Fax: (806) 743-2990 e-mail: [email protected] EDUCATION: Postdoctoral Studies: Department of Tumor Biology, Institute of Medical Science, University of Tokyo, Tokyo, Japan (Advisor: Dr. Yoshikazu Nakamura). Department of Medical Biochemistry and Genetics, College of Medicine, Texas A&M University, College Station, TX (Advisor: Dr. Arthur E. Johnson). Ph.D. in Biochemistry, Russian Academy of Sciences, Pushchino, Moscow Region, Russia. M.S., Biology, Kuban State University, Krasnodar, Russia. FIELDS OF SPECIALIZATION: Biochemistry, Molecular and Cellular Biology. RESEARCH INTERESTS Molecular mechanisms of human diseases. Post-transcriptional regulation of gene expression. Gene silencing and translational repression. mRNA and protein quality control. RNA stability and degradation. siRNAs/miRNAs. Prolactin, CFTR, secretory and membrane proteins. Protein translation, folding and transport. Protein-protein interactions. SCIENTIFIC PUBLICATIONS (see list below) 31 publications (total), including 25 full-length peer-reviewed papers in scientific journals, 5 articles in the Encyclopedia of Molecular Biology (Publisher: John Wiley & Sons, Inc., N.Y.), and chapter in a book. PRESENTATIONS (90 total, see lists below) Results were presented at various international and national meetings, conferences and symposia (58 presentations), as well as during invited or selected talks (32 talks) at different universities and meetings around the world. Andrey L. Karamyshev, Ph.D. RESEARCH EXPERIENCE AND POSITIONS 2016-present Assistant Professor (Tenure track), Department of Cell Biology and Biochemistry!, Texas Tech University Health Sciences Center!, Lubbock, TX. 2009-2016 Assistant Professor (Research track), Department of Physiology, UT Southwestern Medical Center at Dallas, Dallas, TX. -

RNA Interference in the Nucleus: Roles for Small Rnas in Transcription, Epigenetics and Beyond
REVIEWS NON-CODING RNA RNA interference in the nucleus: roles for small RNAs in transcription, epigenetics and beyond Stephane E. Castel1 and Robert A. Martienssen1,2 Abstract | A growing number of functions are emerging for RNA interference (RNAi) in the nucleus, in addition to well-characterized roles in post-transcriptional gene silencing in the cytoplasm. Epigenetic modifications directed by small RNAs have been shown to cause transcriptional repression in plants, fungi and animals. Additionally, increasing evidence indicates that RNAi regulates transcription through interaction with transcriptional machinery. Nuclear small RNAs include small interfering RNAs (siRNAs) and PIWI-interacting RNAs (piRNAs) and are implicated in nuclear processes such as transposon regulation, heterochromatin formation, developmental gene regulation and genome stability. RNA interference Since the discovery that double-stranded RNAs (dsRNAs) have revealed a conserved nuclear role for RNAi in (RNAi). Silencing at both the can robustly silence genes in Caenorhabditis elegans and transcriptional gene silencing (TGS). Because it occurs post-transcriptional and plants, RNA interference (RNAi) has become a new para- in the germ line, TGS can lead to transgenerational transcriptional levels that is digm for understanding gene regulation. The mecha- inheritance in the absence of the initiating RNA, but directed by small RNA molecules. nism is well-conserved across model organisms and it is dependent on endogenously produced small RNA. uses short antisense RNA to inhibit translation or to Such epigenetic inheritance is familiar in plants but has Post-transcriptional gene degrade cytoplasmic mRNA by post-transcriptional gene only recently been described in metazoans. silencing silencing (PTGS). PTGS protects against viral infection, In this Review, we cover the broad range of nuclear (PTGS). -

RNA Stem Structure Governs Coupling of Dicing and Gene Silencing in RNA
RNA stem structure governs coupling of dicing and PNAS PLUS gene silencing in RNA interference Hye Ran Koha,b,1, Amirhossein Ghanbariniakia,c,d, and Sua Myonga,c,d,1 aDepartment of Biophysics, Johns Hopkins University, Baltimore, MD 21218; bDepartment of Chemistry, Chung-Ang University, Seoul 06974, Korea; cInstitute for Genomic Biology, University of Illinois, Urbana, IL 61801; and dCenter for Physics of Living Cells, University of Illinois, Urbana, IL 61801 Edited by Brenda L. Bass, University of Utah School of Medicine, Salt Lake City, UT, and approved October 13, 2017 (received for review June 8, 2017) PremicroRNAs (premiRNAs) possess secondary structures consist- coworkers (20), but its relationship to the silencing efficiency has ing of a loop and a stem with multiple mismatches. Despite the not been tested. While the effect of mismatches between the well-characterized RNAi pathway, how the structural features of guide strand and target mRNA has been extensively studied (21– premiRNA contribute to dicing and subsequent gene-silencing 24), the effect of mismatch between guide and passenger strand efficiency remains unclear. Using single-molecule FISH, we dem- has not been systematically examined. onstrate that cytoplasmic mRNA, but not nuclear mRNA, is reduced We sought to study how different structural features found in during RNAi. The dicing rate and silencing efficiency both increase premiRNAs and presiRNAs modulate overall gene-silencing in a correlated manner as a function of the loop length. In contrast, efficiency by measuring two key steps in the RNAi pathway: (i) mismatches in the stem drastically diminish the silencing efficiency dicing kinetics to measure how quickly premiRNA or presiRNA without impacting the dicing rate. -

(12) Patent Application Publication (10) Pub. No.: US 2004/0086860 A1 Sohail (43) Pub
US 20040O86860A1 (19) United States (12) Patent Application Publication (10) Pub. No.: US 2004/0086860 A1 Sohail (43) Pub. Date: May 6, 2004 (54) METHODS OF PRODUCING RNAS OF Publication Classification DEFINED LENGTH AND SEQUENCE (51) Int. Cl." .............................. C12Q 1/68; C12P 19/34 (76) Inventor: Muhammad Sohail, Marston (GB) (52) U.S. Cl. ............................................... 435/6; 435/91.2 Correspondence Address: MINTZ, LEVIN, COHN, FERRIS, GLOWSKY (57) ABSTRACT AND POPEO, PC. ONE FINANCIAL CENTER Methods of making RNA duplexes and single-stranded BOSTON, MA 02111 (US) RNAS of a desired length and Sequence based on cleavage of RNA molecules at a defined position, most preferably (21) Appl. No.: 10/264,748 with the use of deoxyribozymes. Novel deoxyribozymes capable of cleaving RNAS including a leader Sequence at a (22) Filed: Oct. 4, 2002 Site 3' to the leader Sequence are also described. Patent Application Publication May 6, 2004 Sheet 1 of 2 US 2004/0086860 A1 DNA Oligonucleotides T7 Promoter -TN-- OR 2N-2-N-to y Transcription Products GGGCGAAT-N-UU GGGCGAAT-N-UU w N Deoxyribozyme Cleavage - Q GGGCGAAT -------' Racction GGGCGAAT N-- UU N-UU ssRNA products N-UU Anneal ssRNA UU S-2N- UU siRNA product FIGURE 1: Flowchart summarising the procedure for siRNA synthesis. Patent Application Publication May 6, 2004 Sheet 2 of 2 US 2004/0086860 A1 Full-length transcript 3'-digestion product 5'-digestion product (5'GGGCGAATA) A: Production of single-stranded RNA templates by in vitro transcription and digestion With a deoxyribozyme V 2- 2 V 22inv 22 * 2 &3 S/AS - 88.8x, *...* or as IGFR -- is as 4. -

Switch from Translation Initiation to Elongation Needs Not4 and Not5 Collaboration
bioRxiv preprint doi: https://doi.org/10.1101/850859; this version posted November 22, 2019. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Switch from translation initiation to elongation needs Not4 and Not5 collaboration George E Allen1°, Olesya O Panasenko1°, Zoltan Villanyi1,&, Marina Zagatti1, Benjamin Weiss1, 2 2 1* 5 Christine Polte , Zoya Ignatova , Martine A Collart 1Department of Microbiology and Molecular Medicine, Institute of Genetics and Genomics Geneva, Faculty of Medicine, University of Geneva, Switzerland 2Biochemistry and Molecular Biology, University of Hamburg, Germany °Equally contributing first authors 10 *Correspondence to: [email protected] &Current Address: Dept of Biochemistry and Molecular Biology, University of Szeged Abstract: Not4 and Not5 are crucial components of the Ccr4-Not complex with pivotal functions in mRNA metabolism. Both associate with ribosomes but mechanistic insights on their 15 function remain elusive. Here we determine that Not5 and Not4 synchronously impact translation initiation and Not5 alone alters translation elongation. Deletion of Not5 causes elongation defects in a codon-dependent fashion, increasing and decreasing the ribosome dwelling occupancy at minor and major codons, respectively. This larger difference in codons’ translation velocities alters translation globally and enables kinetically unfavorable processes 20 such as nascent chain deubiquitination to take place. In turn, this leads to abortive translation and favors protein aggregation. These findings highlight the global impact of Not4 and Not5 in controlling the speed of mRNA translation and transition from initiation to elongation. Summary: Not4 and Not5 regulate translation synchronously but distinguishably, facilitating 25 smooth transition from initiation to elongation Results and discussion The Ccr4-Not complex is a global regulator of mRNA metabolism in eukaryotic cells (for review see (1)). -

Expanding the Genetic Code Lei Wang and Peter G
Reviews P. G. Schultz and L. Wang Protein Science Expanding the Genetic Code Lei Wang and Peter G. Schultz* Keywords: amino acids · genetic code · protein chemistry Angewandte Chemie 34 2005 Wiley-VCH Verlag GmbH & Co. KGaA, Weinheim DOI: 10.1002/anie.200460627 Angew. Chem. Int. Ed. 2005, 44,34–66 Angewandte Protein Science Chemie Although chemists can synthesize virtually any small organic molecule, our From the Contents ability to rationally manipulate the structures of proteins is quite limited, despite their involvement in virtually every life process. For most proteins, 1. Introduction 35 modifications are largely restricted to substitutions among the common 20 2. Chemical Approaches 35 amino acids. Herein we describe recent advances that make it possible to add new building blocks to the genetic codes of both prokaryotic and 3. In Vitro Biosynthetic eukaryotic organisms. Over 30 novel amino acids have been genetically Approaches to Protein encoded in response to unique triplet and quadruplet codons including Mutagenesis 39 fluorescent, photoreactive, and redox-active amino acids, glycosylated 4. In Vivo Protein amino acids, and amino acids with keto, azido, acetylenic, and heavy-atom- Mutagenesis 43 containing side chains. By removing the limitations imposed by the existing 20 amino acid code, it should be possible to generate proteins and perhaps 5. An Expanded Code 46 entire organisms with new or enhanced properties. 6. Outlook 61 1. Introduction The genetic codes of all known organisms specify the same functional roles to amino acid residues in proteins. Selectivity 20 amino acid building blocks. These building blocks contain a depends on the number and reactivity (dependent on both limited number of functional groups including carboxylic steric and electronic factors) of a particular amino acid side acids and amides, a thiol and thiol ether, alcohols, basic chain. -
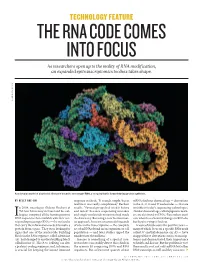
THE RNA CODE COMES INTO FOCUS As Researchers Open up to the Reality of RNA Modification, an Expanded Epitranscriptomics Toolbox Takes Shape
TECHNOLOGY FEATURE THE RNA CODE COMES INTO FOCUS As researchers open up to the reality of RNA modification, an expanded epitranscriptomics toolbox takes shape. LAGUNA DESIGN/SPL LAGUNA A molecular model of a bacterial ribosome bound to messenger RNA, a complex that is formed during protein synthesis. BY KELLY RAE CHI response in check. “It sounds simple, but in mRNAs harbour chemical tags — decorations real life it was really complicated,” Rechavi to the A, C, G and U nucleotides — that are n 2004, oncologist Gideon Rechavi at recalls. “Several groups had tried it before invisible to today’s sequencing technologies. Tel Aviv University in Israel and his col- and failed” because sequencing mistakes (Similar chemical tags, called epigenetic mark- leagues compared all the human genomic and single-nucleotide mutations had made ers, are also found on DNA.) Researchers aren’t IDNA sequences then available with their cor- the data noisy. But using a new bioinformat- sure what these chemical changes in RNA do, responding messenger RNAs — the molecules ics approach, his team uncovered thousands but they’re trying to find out. that carry the information needed to make a of sites in the transcriptome — the complete A wave of studies over the past five years — protein from a gene. They were looking for set of mRNAs found in an organism or cell many of which focus on a specific RNA mark signs that one of the nucleotide building population — and later studies upped the called N6-methyladenosine (m6A) — have blocks in the RNA sequence, called adenosine number into the millions1. -

INTRODUCTION Sirna and Rnai
J Korean Med Sci 2003; 18: 309-18 Copyright The Korean Academy ISSN 1011-8934 of Medical Sciences RNA interference (RNAi) is the sequence-specific gene silencing induced by dou- ble-stranded RNA (dsRNA). Being a highly specific and efficient knockdown tech- nique, RNAi not only provides a powerful tool for functional genomics but also holds Institute of Molecular Biology and Genetics and School of Biological Science, Seoul National a promise for gene therapy. The key player in RNAi is small RNA (~22-nt) termed University, Seoul, Korea siRNA. Small RNAs are involved not only in RNAi but also in basic cellular pro- cesses, such as developmental control and heterochromatin formation. The inter- Received : 19 May 2003 esting biology as well as the remarkable technical value has been drawing wide- Accepted : 23 May 2003 spread attention to this exciting new field. V. Narry Kim, D.Phil. Institute of Molecular Biology and Genetics and School of Biological Science, Seoul National University, San 56-1, Shillim-dong, Gwanak-gu, Seoul 151-742, Korea Key Words : RNA Interference (RNAi); RNA, Small interfering (siRNA); MicroRNAs (miRNA); Small Tel : +82.2-887-8734, Fax : +82.2-875-0907 hairpin RNA (shRNA); mRNA degradation; Translation; Functional genomics; Gene therapy E-mail : [email protected] INTRODUCTION established yet, testing 3-4 candidates are usually sufficient to find effective molecules. Technical expertise accumulated The RNA interference (RNAi) pathway was originally re- in the field of antisense oligonucleotide and ribozyme is now cognized in Caenorhabditis elegans as a response to double- being quickly applied to RNAi, rapidly improving RNAi stranded RNA (dsRNA) leading to sequence-specific gene techniques.