Streptococcus Pneumoniae
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Prevalence of Urinary Tract Infection and Antibiotic Resistance Pattern in Pregnant Women, Najran Region, Saudi Arabia
Vol. 13(26), pp. 407-413, August, 2019 DOI: 10.5897/AJMR2019.9084 Article Number: E3F64FA61643 ISSN: 1996-0808 Copyright ©2019 Author(s) retain the copyright of this article African Journal of Microbiology Research http://www.academicjournals.org/AJMR Full Length Research Paper Prevalence of urinary tract infection and antibiotic resistance pattern in pregnant women, Najran region, Saudi Arabia Ali Mohamed Alshabi1*, Majed Saeed Alshahrani2, Saad Ahmed Alkahtani1 and Mohammad Shabib Akhtar1 1Department of Clinical Pharmacy, College of Pharmacy, Najran University, Najran, Saudi Arabia. 2Department of Obstetics and Gyneocology, Faculty of Medicine, Najran University, Najran, Saudi Arabia. Received 25 February, 2019; Accepted August 5, 2019 Urinary Tract Infection (UTI) is one of the commonest infectious disease in pregnancy, and in pregnancy we have very limited number of antibiotics to treat the UTI. This study was conducted on 151 patients who attended the gynecology clinic during the study period. Nineteen UTI proven cases of UTI were studied for prevalence of microorganism and sensitivity pattern against different antibiotics. Among the bacteria isolated, Escherichia coli (73.68%) and Staphylococcus aureus (10.52%) were the most prevalent Gram negative and Gram positive bacteria respectively. To know the resistance pattern of microorganism we used commercially available discs of different antibiotics. Gram negative bacteria showed more resistance as compared to Gram positive one. It is observed that the most effective antibiotic for Gram negative isolates is Ceftriaxone (87.5%), followed by Amoxicillin + Clavulanic acid (81.25%), Amikacin (75%), Cefuroxime (75%), Cefixime (68.75%) and Mezlocillin (62.5%). For the Gram positive bacteria, Ceftriaxone, Amikacin and Amoxicillin + Clavulanic acid were the most effective antimicrobials (100%). -

Antimicrobial Activity and Antibiotic Resistance of Lactobacillus Delbrueckii Ssp
African Journal of Microbiology Research Vol. 5(6) pp. 675-682, 18 March, 2011 Available online http://www.academicjournals.org/ajmr DOI: 10.5897/AJMR10.835 ISSN 1996-0808 ©2011 Academic Journals Full Length Research Paper Antimicrobial activity and antibiotic resistance of Lactobacillus delbrueckii ssp. bulgaricus and Streptococcus thermophilus strains isolated from Turkish homemade yoghurts Asli Akpinar, Oktay Yerlikaya* and Sevda Kiliç Department of Dairy Technology, Faculty of Agriculture, Ege University, 35100, Bornova – zmir, Turkey. Accepted 1 March, 2011 The aim of this study was to determine the inhibitive effect of 25 Lactobacillus delbruecki ssp. bulgaricus and 16 Streptococcus thermophilus strains isolated from 30 different homemade yoghurts on several pathogen and contaminant bacteria. The antibiotic resistance of these bacteria was also determined. All of Lactobacillus bulgaricus strains exhibited antimicrobial activity against Escherichia coli, whereas all of S. thermophilus strains exhibited the same activity against Klebsiella pneumoniae. None of L. bulgaricus strains were resistant to the polymixin-B, only the OL4 strain has shown resistance to bacitracin. While some strains of S. thermophilus like C6 and SL4 exhibited resistance to novobiocin, SY72, M3, C1M, and F1M were shown to optochin. ET6 and SY73 strains were found to be resistant in both novobiocin and optochin. Key words: Antibiotic resistance, antimicrobial activity, homemade yoghurt, lactic acid bacteria, Lactobacillus delbrueckii ssp. bulgaricus, Streptococcus thermophilus. INTRODUCTION Generally, yoghurt is being perceived as a kind of healthy properties and they have a long history of safe use as food which has a low fat and essential value in terms of starter culture bacteria (Katla et al., 2001). -

Host-Microbe Interactions Defining the Pathogenicity Of
REVIEW published: 10 April 2018 doi: 10.3389/fmicb.2018.00603 The Road to Infection: Host-Microbe Interactions Defining the Pathogenicity of Streptococcus bovis/Streptococcus equinus Complex Members Christoph Jans 1* and Annemarie Boleij 2 1 Laboratory of Food Biotechnology, Institute of Food Nutrition and Health, Department of Health Science and Technology, ETH Zurich, Zurich, Switzerland, 2 Department of Pathology, Radboud Institute for Molecular Life Sciences, Radboudumc, Nijmegen, Netherlands The Streptococcus bovis/Streptococcus equinus complex (SBSEC) comprises several species inhabiting the animal and human gastrointestinal tract (GIT). They match the pathobiont description, are potential zoonotic agents and technological organisms in fermented foods. SBSEC members are associated with multiple diseases in humans and animals including ruminal acidosis, infective endocarditis (IE) and colorectal cancer (CRC). Therefore, this review aims to re-evaluate adhesion and colonization abilities Edited by: of SBSEC members of animal, human and food origin paired with genomic and Sven Hammerschmidt, University of Greifswald, Germany functional host-microbe interaction data on their road from colonization to infection. Reviewed by: SBSEC seem to be a marginal population during GIT symbiosis that can proliferate Konstantinos Papadimitriou, as opportunistic pathogens. Risk factors for human colonization are considered Agricultural University of Athens, Greece living in rural areas and animal-feces contact. Niche adaptation plays a pivotal role Marcelo Gottschalk, where Streptococcus gallolyticus subsp. gallolyticus (SGG) retained the ability to Université de Montréal, Canada proliferate in various environments. Other SBSEC members have undergone genome *Correspondence: reduction and niche-specific gene gain to yield important commensal, pathobiont and Christoph Jans [email protected] technological species. -

Correlation Between Streptococcus Bovis Bacteremia and Density of Cows in Galicia, Northwest of Spain
Infection (2019) 47:399–407 https://doi.org/10.1007/s15010-018-1254-x ORIGINAL PAPER Correlation between Streptococcus bovis bacteremia and density of cows in Galicia, northwest of Spain J. Corredoira1 · E. Miguez2 · L. M. Mateo3 · R. Fernández‑Rodriguez4 · J. F. García‑Rodriguez5 · A. Peréz‑Gonzalez6 · A. Sanjurjo7 · M. V. Pulian8 · R. Rabuñal1 · GESBOGA Received: 8 September 2018 / Accepted: 15 November 2018 / Published online: 29 November 2018 © Springer-Verlag GmbH Germany, part of Springer Nature 2018 Abstract Background There are few data on the epidemiology of infections caused by Streptococcus bovis (Sb). Some studies suggest that both residence in rural areas and contact with livestock could be potential risk factors. Methods We performed a retrospective study for the period 2005–2016 of all cases of bacteremia caused by Sb in Galicia (a region in the northwest of Spain). The association between the incidence rate of Sb bacteremia and the number of cattle by province and district was analyzed. Results 677 cases were included with a median age of 76 years, 69.3% males. The most frequent infections were endocarditis (234 cases, 34.5%), primary bacteremia (213 cases, 31.5%) and biliary infection (119 cases, 17.5%). In 252 patients, colon neoplasms were detected (37.2%). S. gallolyticus subsp. gallolyticus was the predominant species (52.3%). Mortality was 15.5% (105 cases). The annual incidence rate was 20.2 cases/106 inhabitants and was correlated with the density of cattle (p < 0.001), but not with rurality. When comparing the two provinces with a strong predominance of rural population, but with important differences in the number of cattle, such as Orense and Lugo, with 6% and 47.7% of Galician cattle, respectively, the rates were very different: 15.8 and 43.6 cases/106, respectively, with an RR of 2.7 (95% CI, 2.08–3.71). -

Use of the Diagnostic Bacteriology Laboratory: a Practical Review for the Clinician
148 Postgrad Med J 2001;77:148–156 REVIEWS Postgrad Med J: first published as 10.1136/pmj.77.905.148 on 1 March 2001. Downloaded from Use of the diagnostic bacteriology laboratory: a practical review for the clinician W J Steinbach, A K Shetty Lucile Salter Packard Children’s Hospital at EVective utilisation and understanding of the Stanford, Stanford Box 1: Gram stain technique University School of clinical bacteriology laboratory can greatly aid Medicine, 725 Welch in the diagnosis of infectious diseases. Al- (1) Air dry specimen and fix with Road, Palo Alto, though described more than a century ago, the methanol or heat. California, USA 94304, Gram stain remains the most frequently used (2) Add crystal violet stain. USA rapid diagnostic test, and in conjunction with W J Steinbach various biochemical tests is the cornerstone of (3) Rinse with water to wash unbound A K Shetty the clinical laboratory. First described by Dan- dye, add mordant (for example, iodine: 12 potassium iodide). Correspondence to: ish pathologist Christian Gram in 1884 and Dr Steinbach later slightly modified, the Gram stain easily (4) After waiting 30–60 seconds, rinse with [email protected] divides bacteria into two groups, Gram positive water. Submitted 27 March 2000 and Gram negative, on the basis of their cell (5) Add decolorising solvent (ethanol or Accepted 5 June 2000 wall and cell membrane permeability to acetone) to remove unbound dye. Growth on artificial medium Obligate intracellular (6) Counterstain with safranin. Chlamydia Legionella Gram positive bacteria stain blue Coxiella Ehrlichia Rickettsia (retained crystal violet). -

Medical Bacteriology
LECTURE NOTES Degree and Diploma Programs For Environmental Health Students Medical Bacteriology Abilo Tadesse, Meseret Alem University of Gondar In collaboration with the Ethiopia Public Health Training Initiative, The Carter Center, the Ethiopia Ministry of Health, and the Ethiopia Ministry of Education September 2006 Funded under USAID Cooperative Agreement No. 663-A-00-00-0358-00. Produced in collaboration with the Ethiopia Public Health Training Initiative, The Carter Center, the Ethiopia Ministry of Health, and the Ethiopia Ministry of Education. Important Guidelines for Printing and Photocopying Limited permission is granted free of charge to print or photocopy all pages of this publication for educational, not-for-profit use by health care workers, students or faculty. All copies must retain all author credits and copyright notices included in the original document. Under no circumstances is it permissible to sell or distribute on a commercial basis, or to claim authorship of, copies of material reproduced from this publication. ©2006 by Abilo Tadesse, Meseret Alem All rights reserved. Except as expressly provided above, no part of this publication may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without written permission of the author or authors. This material is intended for educational use only by practicing health care workers or students and faculty in a health care field. PREFACE Text book on Medical Bacteriology for Medical Laboratory Technology students are not available as need, so this lecture note will alleviate the acute shortage of text books and reference materials on medical bacteriology. -

MALDI‑TOF MS‑Based Direct‑On‑Target Microdroplet Growth Assay: Latest Developments
EXPERIMENTAL AND THERAPEUTIC MEDICINE 20: 2555-2556, 2020 COMMENT MALDI‑TOF MS‑based direct‑on‑target microdroplet growth assay: Latest developments IOANNIS K. NEONAKIS1 and DEMETRIOS A. SPANDIDOS2 1Department of Clinical Microbiology and Microbial Pathogenesis, University Hospital of Heraklion, 71201 Heraklion; 2Laboratory of Clinical Virology, School of Medicine, University of Crete, 71003 Heraklion, Greece Received June 19, 2020; Accepted July 8, 2020 DOI: 10.3892/etm.2020.8976 Abstract. The matrix-assisted laser desorption-ionization in each droplet spot is known, the MIC can be evaluated and time‑of‑flight mass spectrometry direct‑on‑target microdro- the microorganism can be characterized as susceptible or not. plet growth assay for the rapid susceptibility testing and the With this assay and using a panel of different antibiotics, not detection of the underlying antibiotic resistance mechanisms only can the susceptibility status of an isolate be determined of microbia has been recently introduced. In the present study, but also the underlining resistance mechanisms be revealed. we review the latest developments in the field. In the initial reports, the assay was used for the detection of Recently, the matrix‑assisted laser desorption‑ionization carbapenem nonsusceptibility in Gram‑negative pathogens time‑of‑flight mass spectrometry (MALDI‑TOF MS) both from colonies (1) and directly from positive blood cultures direct‑on‑target microdroplet growth assay (DOT‑MGA) for (BCs) (2). The assay was also used for the detection of two the rapid susceptibility testing and the detection of the under- additional resistance mechanisms among Enterobacterales, lying antibiotic resistance mechanisms of microbia has been namely the extended‑spectrum β‑lactamase (ESBL) produc- introduced (1‑3). -

Transfer of Streptococcus Faecalis and Streptococcus Faecium to the Genus Enterococcus Norn
INTERNATIONALJOURNAL OF SYSTEMATICBACTERIOLOGY, Jan. 1984, p. 31-34 Vol. 34, No. 1 OO20-7713/84/010031-04$02.00/0 Copyright 0 1984, International Union of Microbiological Societies Transfer of Streptococcus faecalis and Streptococcus faecium to the Genus Enterococcus norn. rev. as Enterococcus faecalis comb. nov. and Enterococcus faecium comb. nov. KARL H. SCHLEIFER* AND RENATE KILPPER-BALZ Lehrstuhl fur Mikrobiologie, Technische Universitat Miinchen, D-8000 Miinchen 2, Federal Republic of Germany The results of deoxyribonucleic acid-deoxyribonucleic acid and deoxyribonucleic acid-ribosomal ribonu- cleic acid hybridization studies demonstrated that Streptococcus faecalis and Streptococcus faecium are distantly related to the non-enterococcal streptococci (Streptococcus hovis and Streptococcus equinus) of serological group D and to other streptococci. On the basis of our results and those of previous studies, we propose that S. faecalis and S. faecium be transferred to the genus Enterococcus (ex Thiercelin and Jouhaud) nom. rev. as Enterococcus faecalis (Andrewes and Horder) comb. nov. and Enterococcus faecium (Orla-Jensen) comb. nov., respectively. A description of the genus Enterococcus nom. rev. and emended descriptions of E. faecalis and E. faecium are given. The streptococci belonging to serological group D can be De Ley (4) and were corrected to the value for the reference divided into two physiologically different groups. Strepto- Escherichia coli K-12 DNA. coccus faecalis and Streptococcus faecium were placed in the enterococcus division of the streptococci, whereas RESULTS AND DISCUSSION Streptococcus bovis and Streptococcus equinus were placed in the viridans division by Sherman (21). Kalina proposed (9) The DNA base compositions, serological groups, and that Streptococcus faecalis and Streptococcus faecium peptidoglycan types of the test strains are shown in Table 1. -

Characterization of Antibiotic Resistance Genes in the Species of the Rumen Microbiota
ARTICLE https://doi.org/10.1038/s41467-019-13118-0 OPEN Characterization of antibiotic resistance genes in the species of the rumen microbiota Yasmin Neves Vieira Sabino1, Mateus Ferreira Santana1, Linda Boniface Oyama2, Fernanda Godoy Santos2, Ana Júlia Silva Moreira1, Sharon Ann Huws2* & Hilário Cuquetto Mantovani 1* Infections caused by multidrug resistant bacteria represent a therapeutic challenge both in clinical settings and in livestock production, but the prevalence of antibiotic resistance genes 1234567890():,; among the species of bacteria that colonize the gastrointestinal tract of ruminants is not well characterized. Here, we investigate the resistome of 435 ruminal microbial genomes in silico and confirm representative phenotypes in vitro. We find a high abundance of genes encoding tetracycline resistance and evidence that the tet(W) gene is under positive selective pres- sure. Our findings reveal that tet(W) is located in a novel integrative and conjugative element in several ruminal bacterial genomes. Analyses of rumen microbial metatranscriptomes confirm the expression of the most abundant antibiotic resistance genes. Our data provide insight into antibiotic resistange gene profiles of the main species of ruminal bacteria and reveal the potential role of mobile genetic elements in shaping the resistome of the rumen microbiome, with implications for human and animal health. 1 Departamento de Microbiologia, Universidade Federal de Viçosa, Viçosa, Minas Gerais, Brazil. 2 Institute for Global Food Security, School of Biological -
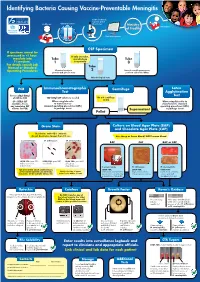
Identifying Bacteria Causing Vaccine-Preventable Meningitis
Identifying Bacteria Causing Vaccine-Preventable Meningitis Sentinel, national, regional, global Health care Clinical specimen laboratories Ministry Vaccination of Health Data management CSF Specimen If specimen cannot be processed in <1 hour, If only one tube, inoculate into Tube microbiology Tube T-I medium. 1 is a priority 3 For details consult Lab Manual or Standard Tube Operating Procedures Chemical analysis: 2 Record overall appearance; protein and glucose tests perform cell count: WBCs Microbiological tests PCR Immunochromatographic Centrifuge Latex Test Agglutination Freeze 250µl-500µl Test of CSF for PCR 100-200µl CSF volume is needed Do not centrifuge if<1ml (if < 500µl CSF When using kits refer When using kits refer to available, freeze to manufacturers manufacturers standard any remaining CSF standard operating procedures (SOPs) operating procedures (SOPs) volume for PCR) in package insert ( Supernatant in package insert Pellet ( Gram Stain Culture on Blood Agar Plate (BAP) and Chocolate Agar Plate (CAP) Decolorize with 95% ethanol; do not decolorize longer than 10 sec. Use sheep or horse blood, NOT human blood S. pneumoniae: H. influenzae: N. meningitidis: BAP CAP BAP or CAP LOOK FOR: gram POS. LOOK FOR: gram NEG. LOOK FOR: gram NEG. diplococci or cocci coccobacilli coffee-bean in short chains shaped diplococci S. pneumoniae H. influenzae N. meningitidis FOR anY otheR GRam stain Results, LOOK FOR: LOOK FOR: LOOK FOR: applY otheR Routine lab methods: Notify clinician of Gram small, grayish, moist (sometimes large, round, colorless- -

Streptococci
STREPTOCOCCI Streptococci are Gram-positive, nonmotile, nonsporeforming, catalase-negative cocci that occur in pairs or chains. Older cultures may lose their Gram-positive character. Most streptococci are facultative anaerobes, and some are obligate (strict) anaerobes. Most require enriched media (blood agar). Streptococci are subdivided into groups by antibodies that recognize surface antigens (Fig. 11). These groups may include one or more species. Serologic grouping is based on antigenic differences in cell wall carbohydrates (groups A to V), in cell wall pili-associated protein, and in the polysaccharide capsule in group B streptococci. Rebecca Lancefield developed the serologic classification scheme in 1933. β-hemolytic strains possess group-specific cell wall antigens, most of which are carbohydrates. These antigens can be detected by immunologic assays and have been useful for the rapid identification of some important streptococcal pathogens. The most important groupable streptococci are A, B and D. Among the groupable streptococci, infectious disease (particularly pharyngitis) is caused by group A. Group A streptococci have a hyaluronic acid capsule. Streptococcus pneumoniae (a major cause of human pneumonia) and Streptococcus mutans and other so-called viridans streptococci (among the causes of dental caries) do not possess group antigen. Streptococcus pneumoniae has a polysaccharide capsule that acts as a virulence factor for the organism; more than 90 different serotypes are known, and these types differ in virulence. Fig. 1 Streptococci - clasiffication. Group A streptococci causes: Strep throat - a sore, red throat, sometimes with white spots on the tonsils Scarlet fever - an illness that follows strep throat. It causes a red rash on the body. -

Optochin Resistance in Streptococcus Pneumoniae: Mechanism, Significance, and Clinical Implications
582 Optochin Resistance in Streptococcus pneumoniae: Mechanism, Significance, and Clinical Implications Andreas Pikis,1,2 Joseph M. Campos,3,4,5,6 1Vaccine and Therapeutic Development Section, Oral Infection William J. Rodriguez,2,4,a and Jerry M. Keith1 and Immunity Branch, National Institute of Dental and Craniofacial Research, National Institutes of Health, Bethesda, Maryland; Departments of 2Infectious Diseases and 3Laboratory Medicine, Children’s National Medical Center, and Departments of 4Pediatrics, 5Pathology, and 6Microbiology/Tropical Medicine, George Washington University Medical Center, Washington, DC Downloaded from https://academic.oup.com/jid/article/184/5/582/808236 by guest on 23 September 2021 Traditionally, Streptococcus pneumoniae is identified in the laboratory by demonstrating susceptibility to optochin. Between 1992 and 1998, 4 pneumococcal isolates exhibiting op- tochin resistance were recovered from patients at Children’s National Medical Center. Three of the 4 isolates consisted of mixed populations of optochin-resistant and -susceptible organ- isms. Both subpopulations had identical antibiograms, serotypes, and restriction fragment profiles. The other isolate was uniformly resistant to optochin. Resistant strains had MICs of optochin 4–30-fold higher than susceptible strains, belonged to different serotypes, and had dissimilar restriction fragment profiles, indicating clonal unrelatedness. Resistance arose from single point mutations in either the a-subunit (W206S) or the c-subunit (G20S, M23I, and A49T) of H+-ATPase. There is speculation of a possible association between exposure to antimalarial drugs and evolution of optochin resistance. a-Hemolytic streptococci resistant to optochin, particularly invasive isolates, should be tested for bile solubility or with an S. pneumoniae DNA probe before identification as viridans streptococci.