Selected Physiological Characteristics of Elite Rowers Measured in the Laboratory and Field and Their Relationship with Performance
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2005 / 2006 the Racewalking Year in Review
2005 / 2006 THE RACEWALKING YEAR IN REVIEW COMPLETE VICTORIAN RESULTS MAJOR INTERNATIONAL RESULTS Tim Erickson 11 November 2006 1 2 Table of Contents AUSTRALIAN UNIVERSITY GAMES, QLD, 27-29 SEPTEMBER 2005......................................................................5 VICTORIAN SCHOOLS U17 – U20 TRACK AND FIELD CHAMPIONSHIPS, SAT 8 OCTOBER 2005...................6 VRWC RACES, ALBERT PARK, SUNDAY 23 OCTOBER 2005...................................................................................7 CHINESE NATIONAL GAMES, NANJING, 17-22 OCTOBER 2005 ..........................................................................10 VICTORIAN ALL SCHOOLS U12-U16 CHAMPIONSHIPS, OLYMPIC PARK, 29 OCTOBER 2005 .....................12 VRWC RACES, ALBERT PARK, SUNDAY 13 NOVEMBER 2005.............................................................................13 PACIFIC SCHOOLS GAMES, MELBOURNE, NOVEMBER 2005..............................................................................16 AUSTRALIAN ALL SCHOOLS CHAMPS, SYDNEY, 8-11 DECEMBER 2005..........................................................19 VRWC RACES, SUNDAY 11 DECEMBER 2005...........................................................................................................23 RON CLARKE CLASSIC MEET, GEELONG, 5000M WALK FOR ELITE MEN, SAT 17 DECEMBER 2005.........26 GRAHAM BRIGGS MEMORIAL TRACK CLASSIC, HOBART, FRI 6 JANUARY 2006..........................................28 NSW 5000M TRACK WALK CHAMPIONSHIPS, SYDNEY, SAT 7 JANUARY 2006...............................................29 -
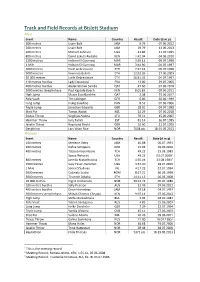
Track and Field Records at Bislett Stadium
Track and Field Records at Bislett Stadium Men Event Name Country Result Date (d.m.y) 100 metres Usain Bolt JAM 9.79 07.06.2012 200 metres Usain Bolt JAM 19.79 13.06.2013 400 metres Michael Johnson USA 43.86 21.07.1995 800 metres David Lekuta Rudisha KEN 1:42.04 04.06.2010 1500 metres Hicham El Guerrouj MAR 3:29.12 09.07.1998 1 Mile Hicham El Guerrouj MAR 3:44.90 04.07.1997 3000 metres Haile Gebrselassie ETH 7:27.42 09.07.1998 5000 metres Kenenisa Bekele ETH 12:52.26 27.06.2003 10 000 metres Haile Gebreslassie ETH 26:31.32 04.07.1997 110 metres hurdles Ladji Doucouré FRA 13.00 29.07.2005 400 metres hurdles Abderrahman Samba QAT 47.60 07.06.2018 3000 metres steeplechase Paul Kipsiele Koech KEN 8:01.83 09.06.2011 High Jump Mutaz Essa Barshim QAT 2.38 15.06.2017 Pole Vault Tim Lobinger GER 6.00 30.06.1999 Long Jump Irving Saladino PAN 8.53 02.06.2006 Triple Jump Jonathan Edwards GBR 18.01 09.07.1998 Shot Put Tomas Walsh NZL 22.29 07.06.2018 Discus Throw Virgilijus Alekna LTU 70.51 15.06.2007 Hammer Throw Jurij Tamm EST 81.14 16.07.1985 Javelin Throw Raymond Hecht GER 92.60 21.07.1995 Decathlon Lars Vikan Rise NOR 7608 pts 18-19.05.2013 Women Event Name Country Result Date (d.m.y) 100 metres Merlene Ottey JAM 10.88 06.07.1991 200 metres Dafne Schippers NED 21.93 09.06.2016 400 metres Tatjana Kocembova TCH 49.23 23.08.1983 Sanya Richards USA 49.23 03.07.20091 800 metres Jarmila Kratochvilova TCH 1:55.04 23.08.19832 1500 metres Suzy Favor-Hamilton USA 3:57.40 28.07.2000 1 Mile Sonia O'Sullivan IRL 4:17.25 22.07.1994 3000 metres Gabriela Szabo -

Athletics SA 2021 State Track and Field Championships
Athletics SA 2021 State Track and Field Championships Final Timetable - as at 25/2/2021 Friday - 26th February Day Time Event Age Group Round Long Jump Triple Jump High Jump Pole Vault Shot Put Discus Javelin Hammer Fri 6.30 PM 3000 metres Walk Under 14 Men & Women FINAL 6.30 PM U17/18/20 Women Fri 3000 metres Walk Under 15 Men & Women FINAL Fri 3000 metres Walk Under 16 Men & Women FINAL Fri 5000 metres Walk Under 17 Men & Women FINAL 6.35 PM U17/18/20 Women Fri 5000 metres Walk Under 18 Men & Women FINAL Fri 5000 metres Walk Under 20 Women FINAL Fri 5000 metres Walk Under Open Women FINAL Fri 5000 metres Walk Under 20 Men FINAL Fri 5000 metres Walk Under Open Men FINAL Fri 5000 metres Walk Over 35 & Over 50 Men & FINAL 6.40 PM Women U14/15/16 Men Fri 6.45 PM 6.45 PM U15/16/U20 Women Fri 6.50 PM 6.50 PM Fri 6.55 PM 6.55 PM Fri 7.00 PM 200 metres Hurdles Under 15 Women FINAL 7.00 PM Fri 200 metres Hurdles Under 16 Women FINAL Fri 7.05 PM 200 metres Hurdles Under 15 Men FINAL 7.05 PM Fri 200 metres Hurdles Under 16 Men FINAL Fri 7.10 PM 200 metres Hurdles Over 35 & Over 50 Men & FINAL 7.10 PM Women Fri 7.15 PM 7.15 PM O35/O50 Women Fri 7.20 PM 400 metres Hurdles Open Men FINAL 7.20 PM Fri 400 metres Hurdles Under 20 Men FINAL 7.25 PM Fri 7:30 PM 400 metres Hurdles Under 17 Men FINAL 7.30 PM Fri 400 metres Hurdles Under 18 Men FINAL Fri 7.35 PM 7.35 PM Open Women Fri 7:40 PM 400 metres Hurdles Under 17 Women FINAL 7.40 PM Seated Fri 400 metres Hurdles Under 18 Women FINAL 7.45 PM U17/18/20 Men U17/18/20 Men Fri 7.50 PM 800 metres Open Men -

YOUTH OLYMPIC GAMES BUENOS AIRES 2018 Quota Places By
APPENDIX 3 YOUTH OLYMPIC GAMES BUENOS AIRES 2018 02-Feb-18 Quota Places by Continent and Event Boys AFRICA AMERICAS ASIA EUROPE OCEANIA totals 100 Metres 5 12 8 6 1 32 200 Metres 3 5 4 11 1 24 400 Metres 5 8 5 5 1 24 800 Metres 8 5 2 8 1 24 1500 Metres 7 3 1 6 1 18 3000 Metres 6 1 5 5 1 18 110m Hurdles 2 8 3 7 2 22 400 Metres Hurdles 3 6 3 3 1 16 2000 Metres Steeplechase 6 2 2 7 1 18 High Jump 1 5 4 5 1 16 Pole Vault 1 1 3 10 1 16 Long Jump 1 6 2 6 1 16 Triple Jump 2 6 4 3 1 16 Shot Put 2 4 2 7 1 16 Discus Throw 1 4 2 8 1 16 Hammer Throw 1 2 2 10 1 16 Javelin Throw 2 3 2 8 1 16 10,000 Metres Race Walk 1 7 3 4 1 16 Totals 57 88 57 119 19 340 Girls AFRICA AMERICAS ASIA EUROPE OCEANIA Totals 100 Metres 3 14 1 13 1 32 200 Metres 1 7 4 11 1 24 400 Metres 3 7 2 11 1 24 800 Metres 5 3 1 14 1 24 1500 Metres 7 2 3 5 1 18 3000 Metres 8 2 3 4 1 18 100m Hurdles 1 8 2 10 1 22 400 Metres Hurdles 1 5 2 7 1 16 2000 Metres Steeplechase 3 3 2 9 1 18 High Jump 1 2 1 11 1 16 Pole Vault 1 3 1 10 1 16 Long Jump 1 5 1 8 1 16 Triple Jump 1 4 2 8 1 16 Shot Put 1 3 1 10 1 16 Discus Throw 2 4 1 8 1 16 Hammer Throw 1 3 1 10 1 16 Javelin Throw 1 5 2 7 1 16 5000 Metres Race Walk 2 5 2 6 1 16 Totals 43 85 32 162 18 340 EVENTS (38) Men’s Events (19) Women’s Events (19) Track Track 100m 100m 200m 200m 400m 400m 800m 800m 1500m* 1500m* 3000m* 3000m* 2000m Steeplechase* 2000m Steeplechase* 110m hurdles 100m hurdles 400m hurdles 400m hurdles 10’000m Race Walk 5’000m Race Walk Field Field High Jump High Jump Pole Vault Pole Vault Long Jump Long Jump Triple Jump Triple Jump Shot Put Shot Put Discus Throw Discus Throw Hammer Throw Hammer Throw Javelin Throw Javelin Throw Cross country* Cross country* Distance (4-6km) Distance (4-6km) Note*: All athletes entered in 1500m, 3000m, 2000m Steeple Chase to compete in Cross Country event. -

Issue No: 363 April/May 2015 Editor: Dave Ainsworth
Issue No: 363 April/May 2015 Editor: Dave Ainsworth PILGRIM's PROGRESS SEEING NO MORE RED Many remember Bill Pilgrim who walked for Borough of Stalwart Cambridge Harriers "Mr Starter" Enfield Harriers (in 1999 they merged with Harringey AC to Dave Maynard has, after 42 years firing his become the formidable Enfield & Harringey AC) and Verlea pistol, hung up his distinctive red jacket and AC (who in March 2002 merged with Hertford & Ware AC to cap. He was recognised when receiving a become Herts Phoenix AC). Bill also completed local runs coveted and much merited 40 years' long and, now in his 60s, this popular Stevenage resident has - service award from the South of England like so many of our readers - progressed into the world of Athletic Association. Dave's still active as a retirement. We wish this former Deputy Headmaster many helper, as seen at Bexley Winter League long and happy years to come; and hope he'll get along to races and elsewhere. We all thank him for see us someday at our events. many years of service with his pistol. GOLD FOR ILFORD PROTAGONIST Ilford's Francisco Reis turned in a truly top class performance at Lee Valley's arena to emphatically win the British Masters' Indoor 3,000 Metres Championship on a steeply banked track. From the "off" he showed a quality field a proverbial clean pair of heels as his gap by the end of lap 2 was such that others could only complete for 2nd place. Francisco broke the tape in 13 minutes and 35 seconds; a personal best time since reaching status, and just over a minute ahead of Olympian Ian Richards. -

Membrane Disrupting Antimicrobial Peptide Dendrimers with Multiple Amino Termini
Electronic Supplementary Material (ESI) for Medicinal Chemistry Communications This journal is © The Royal Society of Chemistry 2011 S1 Supporting Information for: Membrane disrupting antimicrobial peptide dendrimers with multiple amino termini Michaela Stach, a) Noélie Maillard, a) Rameshwar U. Kadam, a) David Kalbermatter, a) Marcel Meury,b) Malcolm G. P. Page, c) Dimitrios Fotiadis, b) Tamis Darbre and Jean-Louis Reymond a) * a) Department of Chemistry and Biochemistry, University of Berne, Freiestrasse 3, CH-3012 Berne Fax: + 41 31 631 80 57; Tel: +41 31 631 43 25, e-mail: [email protected] b) Institute of Biochemistry and Molecular Medicine, University of Berne, Bühlstrasse 28, 3012 Berne, Switzerland c) Basilea Pharmaceutica International Ltd. Table of Contents MATERIAL AND REAGENTS ....................................................................................................................................... 2 COMBINATORIAL LIBRARY ........................................................................................................................................ 3 Library synthesis ................................................................................................................................................. 3 Bead diffusion assay............................................................................................................................................ 3 Sequence determination...................................................................................................................................... -

Security Systems Services World Report
Security Systems Services World Report established in 1974, and a brand since 1981. www.datagroup.org Security Systems Services World Report Database Ref: 56162 This database is updated monthly. Security Systems Services World Report SECURITY SYSTEMS SERVICES WORLD REPORT The Security systems services Report has the following information. The base report has 59 chapters, plus the Excel spreadsheets & Access databases specified. This research provides World Data on Security systems services. The report is available in several Editions and Parts and the contents and cost of each part is shown below. The Client can choose the Edition required; and subsequently any Parts that are required from the After-Sales Service. Contents Description ....................................................................................................................................... 5 REPORT EDITIONS ........................................................................................................................... 6 World Report ....................................................................................................................................... 6 Regional Report ................................................................................................................................... 6 Country Report .................................................................................................................................... 6 Town & Country Report ...................................................................................................................... -

Medicine, Sport and the Body: a Historical Perspective
Carter, Neil. "Science and the Making of the Athletic Body." Medicine, Sport and the Body: A Historical Perspective. London: Bloomsbury Academic, 2012. 81–104. Bloomsbury Collections. Web. 23 Sep. 2021. <http://dx.doi.org/10.5040/9781849662062.ch-004>. Downloaded from Bloomsbury Collections, www.bloomsburycollections.com, 23 September 2021, 13:15 UTC. Copyright © Neil Carter 2012. You may share this work for non-commercial purposes only, provided you give attribution to the copyright holder and the publisher, and provide a link to the Creative Commons licence. 4 Science and the Making of the Athletic Body Introduction At the 1960 Rome Olympics Britain’s Don Thompson won gold in the 50 kilometres walk. Nicknamed Il Topolino (The Little Mouse) by the Italian press, Thompson’s victory was proclaimed heroic and ‘plucky’ in a quintessentially British way. His preparation had been unconventional to say the least. Thompson, who was an insurance clerk, had collapsed in the heat at the Melbourne Games in 1956 and in order to prepare himself for the humidity of Rome he created a steam-room effect in his bathroom using kettles and heaters, and walking up and down continuously on the bathmat. 1 His feat though was set against the growing rivalry in international sport between America and the USSR where greater resources in terms of coaching and sports science were being dedicated to the preparation of their athletes for the Olympic arena. By contrast, Thompson’s preparation refl ected British sport’s amateur tradition. This chapter charts the evolution of athletes’ training methods since the early nineteenth century. -

Standards Scheme 2019-2020
Under 15 Girls Event Grade 1 Grade 2 Grade 3 Grade 4 Under100 15metres Girls 12.7 sec * 12.9 sec * 13.2 sec 13.5 sec Event200 metres Grade26.2 1 sec * Grade26.6 2 sec * Grade27.3 sec3 Grade28.0 sec 4 300 metres ^ 42.6 sec 43.5 sec 44.4 sec 45.9 sec 100 metres800 metres 12.72 sec min 19.4 sec 12.92 secmin 22.3 sec 13.12 minsec * 25.6 sec * 13.52 min sec 31.2 sec # 200 metres1,500 metres 26.34 sec min 48.7 sec # 26.74 secmin 53.9 sec * 27.25min sec 02.0 sec # 28.05 min sec 15.5 sec# 300 metres3,000 metres 42.310 sec min * 22.0 sec #43.2 10 sec min * 41.5 sec #44.2 11 sec min * 02.5 sec # 45.711 secmin 36.0 sec # 800 metres75 metres Hurdles 2 min11.9 19.3 sec sec * * 2 min12.3 22.1 sec sec * * 2 min12.7 25.6 sec sec * # 2 min13.4 31.3 sec *sec # 1,500High metres Jump 4 min1.57 49.0 metres sec * * 4 min1.53 54.5 metres sec * * 5 min1.47 01.5 metres sec * # 5 min1.40 13.5 metres sec * 3,000Pole metres Vault 10 min2.95 27.0 metres sec * # 10 min2.80 36.0 metres sec *# 11 2.50min 02.0metres sec # # 112.15 min metres#34.5 sec * Long Jump 5.05 metres* 4.90 metres * 4.70 metres 4.45 metres 75 metres Hurdles 11.8 sec * 12.2 sec 12.6 sec 13.4 sec Shot (3K) ^ 10.60 metres 9.85 metres 9.10 metres 8.30 metres High Jump 1.58 metres * 1.54 metres 1.50 metres * 1.41 metres Discus 28.30 metres * 25.65 metres * 23.00 metres * 19.50 metres # Pole Vault 2.90 metres # 2.75 metres # 2.50 metres 2.20 metres Hammer 42.60 metres 37.80 metres # 31.10 metres # 24.50 metres Long Jump 5.10 metres * 4.95 metres * 4.75 metres * 4.45 metres # Javelin (500g) ^ 33.10 metres -

The Training of Women Marathon Runners
fiiggl •£' by IAAF 8:4:61-70, 1993 The training of women marathon runners by Jose Manuel Ballesteros (^C The author outlines some general 1 Introduction iraining principles for the Maraihon and discusses importanl factors which women Women have been running the Maraihon marathon runners must consider. He also seriously only since the late 1970's. Since that describes the ba.sic training methods which, he lime their Increasing interest and willingness believes, will help women perform lo their to compele in ihis evenl have led to a consid full potential in this event. erable improvemenls lo results. However, this article will show that women have the V physical and mental ability to run the marathon distance fasler than they do at pre senl. 1 will cover the general training principles for the Maraihon and discuss some imporlani factors which women marathoners must take into account. I will also describe the basic training methods which I believe should enable women to perform to their full potential. 2 Performance potential The highly specific nalure of modern athlet ics is now apparent In the Marathon. This race has now become a specialised event, as is shown by the fact that only three women were ranked in the World Top 20 both for 10,000 metres and the Maraihon in 1992. It has ceased to be the refuge of 10,000 metres track runners of limiled poiential or alhleies nearing the end of Ihelr careers, Jose Manuel Ballesteros is a member ofthe The evenl is also being taken up at a much NSA AdvLsory Ediiorial Board. -

ATHLETICS AUSTRALIA AUSTRALIAN RECORDS AS at 18Th July 2019
ATHLETICS AUSTRALIA AUSTRALIAN RECORDS AS AT 18th July 2019. MEN EVENT PERF. DATE VENUE NAME STATE 100 METRES Aust Allcomers 9.86999999999999923-Sep-00 Sydney Maurice Greene USA Aust National 9.93 5-May-03 Mita, JPN Patrick Johnson ACT Aust U20 10.15 1-Jul-18 Finland Jake Doran QLD Aust U18 10.38 11-Dec-15 Perth Jack Hale TAS Aust U16 10.67 7-Dec-13 Townsville Jordon Shelley NSW 200 METRES Aust Allcomers 19.92 25-Feb-99 Melbourne Frankie Fredericks NAM Aust National 20.06 16-Oct-68 Mexico City Peter Norman VIC Aust U20 20.48 3-Feb-85 Brisbane Fred Martin NSW Aust U18 20.90 6-Nov-82 Sydney Darren Clark NSW 20.90 19-Aug-89 Saga Paul Greene NSW 20.90 24-Sep-17 Townsville Zane Branco QLD Aust U16 21.44 8-Dec-13 Townsville Jordon Shelley NSW 400 METRES Aust Allcomers 43.84 25-Sep-00 Sydney Michael Johnson USA Aust National 44.38 26-Sep-88 Seoul Darren Clark NSW Aust U20 44.75 8-Aug-84 Los Angeles Darren Clark NSW Aust U18 45.96 19-Aug-89 Saga Paul Greene NSW Aust U16 48.12 7-Dec-88 Sydney Paul Greene NSW 800 METRES Aust Allcomers 1:43.15 4-Mar-10 Melbourne David Rudisha KEN Aust National 1:44.21 20-Jul-18 Monaco Joseph Deng VIC Aust U20 1:45.91 28-Jul-95 Lindau Paul Byrne VIC Aust U18 1.47.24 11-Dec-93 Canberra Paul Byrne VIC Aust U16 1:51.41 11-Dec-88 Canberra Mark Holcombe VIC 1000 METRES Aust Allcomers 2:17.67 14-Jan-88 Melbourne Michael Hillardt QLD Aust National 2:16.09 17-Jun-14 Ostrava Jeffrey Riseley VIC Aust U20 2:20.5 6-Jan-94 Melbourne Paul Byrne VIC Aust U18 2:23.1 7-Jan-93 Melbourne Michael Power VIC Aust U16 2:28.7 7-Jan-92 Melbourne -

Rik Merchie Table of Contents ULTRA TOUR MONTE ROSA 6 MY PUB 8 HARDROCK HUNDRED MILE ENDURANCE RUN 10 3, 2, 1
THE CHALLENGE legendary trails 12 you should run Rik Merchie Table of contents ULTRA TOUR MONTE ROSA 6 MY PUB 8 HARDROCK HUNDRED MILE ENDURANCE RUN 10 3, 2, 1... GO! 12 TOR DES GÉANTS ® 16 BACKPACK 18 TROMSØ SKYRACE 20 FOOTBALL 24 EL CRUCE COLUMBIA 28 JOTIE 36 SALOMON GLEN COE SKYLINE 40 DAD 42 BARKLEY MARATHONS 44 BELGICANO 50 THE NORTH FACE ® LAVAREDO ULTRA TRAIL 54 SCREEN WORKER 58 MARATHON DU MONT-BLANC 62 INFLAMMATION 64 LEADVILLE TRAIL 100 RUN 68 SEX 76 ZEGAMA-AIZKORRI 80 POOP 86 WESTERN STATES 100-MILE ENDURANCE RUN 90 JUNGLE 92 SALOMON CAPPADOCIA ULTRA-TRAIL ® 98 MILESTONE 106 TARAWERA ULTRAMARATHON 110 GOD 116 EVEREST TRAIL RACE ® 120 STORM 122 VIBRAM HONG KONG 100 ULTRA TRAIL RACE 128 FAR WEST 136 ULTRA-TRAIL CAPE TOWN ® 140 PREGNANT 148 MATTERHORN ULTRAKS 152 ULTRA-TRAIL DU MONT-BLANC ® 154 SUMMER 160 MUSTANG MOUNTAIN TRAIL RACE 164 SHEPHERD 170 FESTIVAL DES TEMPLIERS 174 PARTY 176 MARATHON DES SABLES 182 WOLF 184 OTTER AFRICAN TRAIL RUN 190 TITLE 192 WORD OF THANKS 199 BIBLIOGRAPHY 201 Italy & Switzerland ULTRA TOUR MONTE ROSA 170km | 105mi • D+ 11,300m | 37,073ft ULTRA & 4-STAGE RACE 100km | 62mi • D+ 6,100 m | 20,013ft “While I was running these trails around Monte Rosa, my training ground for the UTMB®, I thought this is the race I would really like to run. It didn’t exist so I’ve created it for you to enjoy.” Lizzy Hawker 6 MY PUB ‘If dogs run free, then why not we?' Bob Dylan t’s daybreak when my train worms out My publisher sent me off with the message of the station of the Swiss town of that, for him, twelve races were more than St.