Distribution and Abundance of Submerged Aquatic Vegetation in the Chesapeake Bay: a Scientific Summary
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Title 26 Department of the Environment, Subtitle 08 Water
Presented below are water quality standards that are in effect for Clean Water Act purposes. EPA is posting these standards as a convenience to users and has made a reasonable effort to assure their accuracy. Additionally, EPA has made a reasonable effort to identify parts of the standards that are not approved, disapproved, or are otherwise not in effect for Clean Water Act purposes. Title 26 DEPARTMENT OF THE ENVIRONMENT Subtitle 08 WATER POLLUTION Chapters 01-10 2 26.08.01.00 Title 26 DEPARTMENT OF THE ENVIRONMENT Subtitle 08 WATER POLLUTION Chapter 01 General Authority: Environment Article, §§9-313—9-316, 9-319, 9-320, 9-325, 9-327, and 9-328, Annotated Code of Maryland 3 26.08.01.01 .01 Definitions. A. General. (1) The following definitions describe the meaning of terms used in the water quality and water pollution control regulations of the Department of the Environment (COMAR 26.08.01—26.08.04). (2) The terms "discharge", "discharge permit", "disposal system", "effluent limitation", "industrial user", "national pollutant discharge elimination system", "person", "pollutant", "pollution", "publicly owned treatment works", and "waters of this State" are defined in the Environment Article, §§1-101, 9-101, and 9-301, Annotated Code of Maryland. The definitions for these terms are provided below as a convenience, but persons affected by the Department's water quality and water pollution control regulations should be aware that these definitions are subject to amendment by the General Assembly. B. Terms Defined. (1) "Acute toxicity" means the capacity or potential of a substance to cause the onset of deleterious effects in living organisms over a short-term exposure as determined by the Department. -

News Release Address: Email and Homepage: U.S
News Release Address: Email and Homepage: U.S. Department of the Interior Maryland-Delaware-D.C. District [email protected] U.S. Geological Survey 8987 Yellow Brick Road http://md.water.usgs.gov/ Baltimore, MD 21237 Release: Contact: Phone: Fax: January 4, 2002 Wendy S. McPherson (410) 238-4255 (410) 238-4210 Below Normal Rainfall and Warm Temperatures Lead to Record Low Water Levels in December Three months of above normal temperatures and four months of below normal rainfall have led to record low monthly streamflow and ground-water levels, according to hydrologists at the U.S. Geological Survey (USGS) in Baltimore, Maryland. Streamflow was below normal at 94 percent of the real-time USGS gaging stations and 83 percent of the USGS observation wells across Maryland and Delaware in December. Record low streamflow levels for December were set at Winters Run and Pocomoke River. Streamflow levels at Deer Creek and Winters Run in Harford County have frequently set new record daily lows for the last four months (see real-time graphs at http://md.water.usgs.gov/realtime/). Streamflow was also significantly below normal at Antietam Creek, Choptank River, Conococheague Creek, Nassawango Creek, Patapsco River, Gunpowder River, Patuxent River, Piscataway Creek, Monocacy River, and Potomac River in Maryland, and Christina River, St. Jones River, and White Clay Creek in Delaware. The monthly streamflow in the Potomac River near Washington, D.C. was 82 percent below normal in December and 54 percent below normal for 2001. Streamflow entering the Chesapeake Bay averaged 23.7 bgd (billion gallons per day), which is 54 percent below the long-term average for December. -

Flood Insurance Study
FLOOD INSURANCE STUDY CECIL COUNTY, MARYLAND AND INCORPORATED AREAS Cecil County Community Community Name Number ↓ CECIL COUNTY (UNINCORPORATED AREAS) 240019 *CECILTON, TOWN OF 240020 CHARLESTOWN, TOWN OF 240021 CHESAPEAKE CITY, TOWN OF 240099 ELKTON, TOWN OF 240022 NORTH EAST, TOWN OF 240023 PERRYVILLE, TOWN OF 240024 PORT DEPOSIT, TOWN OF 240025 RISING SUN, TOWN OF 240158 *No Special Flood Hazard Areas Identified Revised: May 4, 2015 Federal Emergency Management Agency FLOOD INSURANCE STUDY NUMBER 24015CV000B NOTICE TO FLOOD INSURANCE STUDY USERS Communities participating in the National Flood Insurance Program (NFIP) have established repositories of flood hazard data for floodplain management and flood insurance purposes. This Flood Insurance Study (FIS) report may not contain all data available within the Community Map Repository. Please contact the Community Map Repository for any additional data. Part or all of this FIS may be revised and republished at any time. In addition, part of the FIS may be revised by the Letter of Map Revision (LOMR) process, which does not involve republication or redistribution of the FIS. It is, therefore, the responsibility of the user to consult with community officials and to check the community repository to obtain the most current FIS components. Initial Countywide FIS Effective Date: July 8, 2013 Revised Countywide FIS Effective Date: May 4, 2015 TABLE OF CONTENTS Page 1.0 INTRODUCTION ............................................................................................................. -

MARK-RECAPTURE ASSESSMENT of the RECREATIONAL BLUE CRAB (Callinectes Sapidus) HARVEST in CHESAPEAKE BAY, MARYLAND
ABSTRACT Title of Thesis: MARK-RECAPTURE ASSESSMENT OF THE RECREATIONAL BLUE CRAB (Callinectes sapidus) HARVEST IN CHESAPEAKE BAY, MARYLAND Robert Francis Semmler, Master of Science, 2016 Directed By: Professor, Marjorie Reaka, Marine Estuarine Environmental Science In Maryland, commercial blue crab (Callinectes sapidus) harvests are monitored through mandatory, annual harvest reporting, but no annual monitoring exists for recreational fishers. This study used a large-scale mark-recapture program to assess relative exploitation between the recreational and commercial fishing sectors in 15 harvest reporting areas of Maryland, then incorporated movement information and extrapolated reported commercial harvest data to generate statewide estimates of recreational harvest. Results indicate spatial variation in recreational fishing, with a majority of recreational harvests coming from tributaries of the Western Shore and the Wye and Miles Rivers on the Eastern Shore. Statewide, recreational harvest has remained approximately 8% as large as commercial harvest despite management changes in 2008, and remains a larger proportion (12.8%) of male commercial harvest. In addition, this study provides detailed spatial information on recreational harvest and the first information on rates of exchange of male crabs among harvest reporting areas. MARK-RECAPTURE ASSESSMENT OF THE RECREATIONAL BLUE CRAB (Callinectes sapidus) HARVEST IN CHESAPEAKE BAY, MARYLAND By Robert Francis Semmler Thesis submitted to the Faculty of the Graduate School of the University of Maryland, College Park, in partial fulfillment of the requirements for the degree of Master of Science, 2016 Advisory Committee: Professor Anson H. Hines, Co-Chair Professor Marjorie L. Reaka, Co-Chair Professor Elizabeth W. North Dr. Matthew B. Ogburn © Copyright by Robert Francis Semmler 2016 Acknowledgements I would like to thank Dr. -
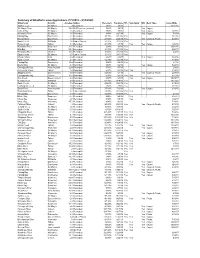
Summary of Lease Applications 9-23-20.Xlsx
Summary of Shellfish Lease Applications (1/1/2015 - 9/23/2020) Waterbody County AcreageStatus Received CompleteTFL Sanctuary WC Gear Type IssuedDate Smith Creek St. Mary's 3 Recorded 1/6/15 1/6/15 11/21/16 St. Marys River St. Mary's 16.2 GISRescreen (revised) 1/6/15 1/6/15 Yes Cages Calvert Bay St. Mary's 2.5Recorded 1/6/15 1/6/15 YesCages 2/28/17 Wicomico River St. Mary's 4.5Recorded 1/8/15 1/27/15 YesCages 5/8/19 Fishing Bay Dorchester 6.1 Recorded 1/12/15 1/12/15 Yes 11/2/15 Honga River Dorchester 14Recorded 2/10/15 2/26/15Yes YesCages & Floats 6/27/18 Smith Creek St Mary's 2.6 Under Protest 2/12/15 2/12/15 Yes Harris Creek Talbot 4.1Recorded 2/19/15 4/7/15 Yes YesCages 4/28/16 Wicomico River Somerset 26.7Recorded 3/3/15 3/3/15Yes 10/20/16 Ellis Bay Wicomico 69.9Recorded 3/19/15 3/19/15Yes 9/20/17 Wicomico River Charles 13.8Recorded 3/30/15 3/30/15Yes 2/4/16 Smith Creek St. Mary's 1.7 Under Protest 3/31/15 3/31/15 Yes Chester River Kent 4.9Recorded 4/6/15 4/9/15 YesCages 8/23/16 Smith Creek St. Mary's 2.1 Recorded 4/23/15 4/23/15 Yes 9/19/16 Fishing Bay Dorchester 12.4Recorded 5/4/15 6/4/15Yes 6/1/16 Breton Bay St. -

Choptank Tributary Summary: a Summary of Trends in Tidal Water Quality and Associated Factors, 1985-2018
Choptank Tributary Summary: A summary of trends in tidal water quality and associated factors, 1985-2018. June 7, 2021 Prepared for the Chesapeake Bay Program (CBP) Partnership by the CBP Integrated Trends Analysis Team (ITAT) This tributary summary is a living document in draft form and has not gone through a formal peer review process. We are grateful for contributions to the development of these materials from the following individuals: Jeni Keisman, Rebecca Murphy, Olivia Devereux, Jimmy Webber, Qian Zhang, Meghan Petenbrink, Tom Butler, Zhaoying Wei, Jon Harcum, Renee Karrh, Mike Lane, and Elgin Perry. 1 Contents 1. Purpose and Scope .................................................................................................................................... 3 2. Location ..................................................................................................................................................... 4 2.1 Watershed Physiography .................................................................................................................... 4 2.2 Land Use .............................................................................................................................................. 6 Land Use ................................................................................................................................................ 6 2.3 Tidal Waters and Stations ................................................................................................................... 8 3. Tidal -

Maryland Stream Waders 10 Year Report
MARYLAND STREAM WADERS TEN YEAR (2000-2009) REPORT October 2012 Maryland Stream Waders Ten Year (2000-2009) Report Prepared for: Maryland Department of Natural Resources Monitoring and Non-tidal Assessment Division 580 Taylor Avenue; C-2 Annapolis, Maryland 21401 1-877-620-8DNR (x8623) [email protected] Prepared by: Daniel Boward1 Sara Weglein1 Erik W. Leppo2 1 Maryland Department of Natural Resources Monitoring and Non-tidal Assessment Division 580 Taylor Avenue; C-2 Annapolis, Maryland 21401 2 Tetra Tech, Inc. Center for Ecological Studies 400 Red Brook Boulevard, Suite 200 Owings Mills, Maryland 21117 October 2012 This page intentionally blank. Foreword This document reports on the firstt en years (2000-2009) of sampling and results for the Maryland Stream Waders (MSW) statewide volunteer stream monitoring program managed by the Maryland Department of Natural Resources’ (DNR) Monitoring and Non-tidal Assessment Division (MANTA). Stream Waders data are intended to supplementt hose collected for the Maryland Biological Stream Survey (MBSS) by DNR and University of Maryland biologists. This report provides an overview oft he Program and summarizes results from the firstt en years of sampling. Acknowledgments We wish to acknowledge, first and foremost, the dedicated volunteers who collected data for this report (Appendix A): Thanks also to the following individuals for helping to make the Program a success. • The DNR Benthic Macroinvertebrate Lab staffof Neal Dziepak, Ellen Friedman, and Kerry Tebbs, for their countless hours in -

Summary of Decisions Regarding Nutrient and Sediment Load Allocations and New Submerged Aquatic Vegetation (SAV) Restoration Goals
To: Principal Staff Committee Members and Representatives of Chesapeake Bay “Headwater” States From: W. Tayloe Murphy, Jr., Chair Chesapeake Bay Program Principals’ Staff Committee Subject: Summary of Decisions Regarding Nutrient and Sediment Load Allocations and New Submerged Aquatic Vegetation (SAV) Restoration Goals For the past twenty years, the Chesapeake Bay partners have been committed to achieving and maintaining water quality conditions necessary to support living resources throughout the Chesapeake Bay ecosystem. In the past month, Chesapeake Bay Program partners (Maryland, Virginia, Pennsylvania, the District of Columbia, the Environmental Protection Agency and the Chesapeake Bay Commission) have expanded our efforts by working with the headwater states of Delaware, West Virginia and New York to adopt new cap load allocations for nitrogen, phosphorus and sediment. Using the best scientific information available, Bay Program partners have agreed to allocations that are intended to meet the needs of the plants and animals that call the Chesapeake home. The allocations will serve as a basis for each state’s tributary strategies that, when completed by April 2004, will describe local implementation actions necessary to meet the Chesapeake 2000 nutrient and sediment loading goals by 2010. This memorandum summarizes the important, comprehensive agreements made by Bay watershed partners with regard to cap load allocations for nitrogen, phosphorus and sediments, as well as new baywide and local SAV restoration goals. Nutrient Allocations Excessive nutrients in the Chesapeake Bay and its tidal tributaries promote undesirable algal growth, and thereby, prohibit light from reaching underwater bay grasses (submerged aquatic vegetation or SAV) and depress the dissolved oxygen levels of the deeper waters of the Bay. -

Maryland's Lower Choptank River Cultural Resource Inventory
Maryland’s Lower Choptank River Cultural Resource Inventory by Ralph E. Eshelman and Carl W. Scheffel, Jr. “So long as the tides shall ebb and flow in Choptank River.” From Philemon Downes will, Hillsboro, circa 1796 U.S. Geological Survey Quadrangle 7.5 Minute Topographic maps covering the Lower Choptank River (below Caroline County) include: Cambridge (1988), Church Creek (1982), East New Market (1988), Oxford (1988), Preston (1988), Sharp Island (1974R), Tilghman (1988), and Trappe (1988). Introduction The Choptank River is Maryland’s longest river of the Eastern Shore. The Choptank River was ranked as one of four Category One rivers (rivers and related corridors which possess a composite resource value with greater than State signific ance) by the Maryland Rivers Study Wild and Scenic Rivers Program in 1985. It has been stated that “no river in the Chesapeake region has done more to shape the character and society of the Eastern Shore than the Choptank.” It has been called “the noblest watercourse on the Eastern Shore.” Name origin: “Chaptanck” is probably a composition of Algonquian words meaning “it flows back strongly,” referring to the river’s tidal changes1 Geological Change and Flooded Valleys The Choptank River is the largest tributary of the Chesapeake Bay on the eastern shore and is therefore part of the largest estuary in North America. This Bay and all its tributaries were once non-tidal fresh water rivers and streams during the last ice age (15,000 years ago) when sea level was over 300 feet below present. As climate warmed and glaciers melted northward sea level rose, and the Choptank valley and Susquehanna valley became flooded. -

2021 Regular Session
HOUSE BILL 843 M2 1lr2482 By: Delegate Howard Introduced and read first time: January 29, 2021 Assigned to: Environment and Transportation A BILL ENTITLED 1 AN ACT concerning 2 Anne Arundel County – Natural Resources – Fishing With a Haul Seine 3 FOR the purpose of repealing certain provisions of law that restrict the length of a haul 4 seine that a person may use in certain waters of Anne Arundel County; repealing a 5 certain penalty for a violation of certain provisions of law that restrict the length of 6 a haul seine that a person may use in certain waters of Anne Arundel County; 7 authorizing a person to use a haul seine to fish in the waters of Anne Arundel County 8 except under certain circumstances; making technical, stylistic, and clarifying 9 changes; and generally relating to fishing with a haul seine in Anne Arundel County. 10 BY repealing and reenacting, without amendments, 11 Article – Natural Resources 12 Section 4–713 13 Annotated Code of Maryland 14 (2018 Replacement Volume and 2020 Supplement) 15 BY repealing and reenacting, with amendments, 16 Article – Natural Resources 17 Section 4–714 18 Annotated Code of Maryland 19 (2018 Replacement Volume and 2020 Supplement) 20 SECTION 1. BE IT ENACTED BY THE GENERAL ASSEMBLY OF MARYLAND, 21 That the Laws of Maryland read as follows: 22 Article – Natural Resources 23 4–713. 24 (a) A person who fishes with haul seine equipment shall possess a tidal fish 25 license to catch finfish. EXPLANATION: CAPITALS INDICATE MATTER ADDED TO EXISTING LAW. [Brackets] indicate matter deleted from existing law. -

Shorerivers-Advocate-Spring-2020
ShoreRivers Advocate CHESTER RIVER | CHOPTANK RIVER MILES & WYE RIVERS | SASSAFRAS RIVER SPRING 2020 Combatting climate change Improving Farm Resiliency through Soil Health & Water Management ShoreRivers Supports LOCAL GREEN SCHOOLS 2019 Donor Appreciation and more… LETTER FROM THE EXECUTIVE DIRECTOR The cover of this edition of the Advocate newsletter footprint, build resiliency into our natural systems, and presents an iconic image of the impacts of climate plan for the future. change on the Eastern Shore. Unfortunately, as you’ll Advocacy: We are promoting legislation to require all discover in the following articles, it is neither a rare development in flood-prone areas to take climate change occurrence, nor is it the only type of impact we are impacts into account. We are recommending regulatory experiencing. The ecological effects on our region of a changes to address increased pollution from construction changing climate include drowned forests, higher tides, site runoff associated with increasingly stronger storms. salty farm fields, and fresher water flowing in our brackish rivers. This translates into Restoration: Restoring our climate crisis: costly the landscape to its infrastructure fixes, natural form protects our canceled events and loss of communities, stores carbon tourism, lower home values, from the atmosphere, flooded main streets, more and builds resiliency in polluted rivers, dangerous the face of a changing swimming conditions, loss future. ShoreRivers excels of productive farmland, and in projects including I could go on. The fact is, a wetland and stream changing climate with restoration, innovative higher temperatures, water management on stronger and more farmland, streamside buffer intense storms, and rising installation, and planting seas increases pollution in our rivers. -

Defining the Nanticoke Indigenous Cultural Landscape
Indigenous Cultural Landscapes Study for the Captain John Smith Chesapeake National Historic Trail: Nanticoke River Watershed December 2013 Kristin M. Sullivan, M.A.A. - Co-Principal Investigator Erve Chambers, Ph.D. - Principal Investigator Ennis Barbery, M.A.A. - Research Assistant Prepared under cooperative agreement with The University of Maryland College Park, MD and The National Park Service Chesapeake Bay Annapolis, MD EXECUTIVE SUMMARY The Nanticoke River watershed indigenous cultural landscape study area is home to well over 100 sites, landscapes, and waterways meaningful to the history and present-day lives of the Nanticoke people. This report provides background and evidence for the inclusion of many of these locations within a high-probability indigenous cultural landscape boundary—a focus area provided to the National Park Service Chesapeake Bay and the Captain John Smith Chesapeake National Historic Trail Advisory Council for the purposes of future conservation and interpretation as an indigenous cultural landscape, and to satisfy the Identification and Mapping portion of the Chesapeake Watershed Cooperative Ecosystems Studies Unit Cooperative Agreement between the National Park Service and the University of Maryland, College Park. Herein we define indigenous cultural landscapes as areas that reflect “the contexts of the American Indian peoples in the Nanticoke River area and their interaction with the landscape.” The identification of indigenous cultural landscapes “ includes both cultural and natural resources and the wildlife therein associated with historic lifestyle and settlement patterns and exhibiting the cultural or esthetic values of American Indian peoples,” which fall under the purview of the National Park Service and its partner organizations for the purposes of conservation and development of recreation and interpretation (National Park Service 2010:4.22).