Some Aspects of the Biology and Host-Parasite Interactions of Isospora Spp
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Two New Species of Eimeria and Three New Species of Isospora (Apicomplexa, Eimeriidae) from Brazilian Mammals and Birds
Bull. Mus. nain. Hist. nat., Paris, 4' sér., 11, 1989, section A, n° 2 : 349-365. Two new species of Eimeria and three new species of Isospora (Apicomplexa, Eimeriidae) from Brazilian mammals and birds by R. LAINSON and J. J. SHAW Abstract. — Thirteen " four-eyed opossums ", Philander o. opossum, from Para State, north Brazil, were examined for coccidial oocysts in faecal samples. Five were infected with an eimerian, considered a new species, 2 with an isosporan which is possibly /. boughtoni Volk, and 2 with both thèse parasites. Oocysts of Eimeria philanderi n. sp., are spherical to subspherical and average 23.50 x 22.38 (21.25- 27.50 x 18.75-25.00) (xm : single polar body : no oocyst residuum. Oocyst wall 1.88 [ira, with mamillated surface and composed of two striated layers, the inner brown-yellow and the outer colourless : no micropyle. Sporocysts average 11.35 x 8.13 (10.00-12.00 x 7.50-8.75) (xm, oval to ellipsoidal, with prominent nipple-like Stieda body : residuum bulky, compact or scattered between two recurved sporozoites. Oocysts of the Isospora sp., close to /. boughtoni initially sub-spherical, 17.92 x 16.53 (16.25- 20.00 x 13.75-18.75) (xm : latterly deformed by collapse of the délicate, colourless wall : no micropyle, oocyst residuum or polar body. Sporocysts 13.35 x 9.88 (12.50-13.75 x 8.75-11.25) (xm, ellipsoidal, with no Stieda body. Two of 5 " woolly opossums ", Caluromys p. philander, were infected with an eimerian considered as a new species, Eimeria caluromydis n. -

Some Parasites of the Common Crow, Corvus Brachyrhynchos Brehm, from Ohio1' 2
SOME PARASITES OF THE COMMON CROW, CORVUS BRACHYRHYNCHOS BREHM, FROM OHIO1' 2 JOSEPH JONES, JR. Biology Department, Saint Augustine's College, Raleigh, North Carolina ABSTRACT Thirty-one species of parasites were taken from 339 common crows over a twenty- month period in Ohio. Of these, nine are new host records: the cestodes Orthoskrjabinia rostellata and Hymenolepis serpentulus; the nematodes Physocephalus sexalatus, Splendido- filaria quiscali, and Splendidofilaria flexivaginalis; and the arachnids Laminosioptes hymenop- terus, Syringophilus bipectinatus, Analges corvinus, and Gabucinia delibata. Twelve parasites not previously reported from the crow in Ohio were also recognized. Two tables, one showing the incidence and intensity of parasitism in the common crow in Ohio, the other listing previous published and unpublished records of common crow parasites, are included. INTRODUCTION Although the crow is of common and widespread occurrence east of the Rockies, no comprehensive, year-round study of parasitism in this bird has been reported. Surveys of parasites of common crows, collected for the most part during the winter season, have been made by Ward (1934), Morgan and Waller (1941), and Daly (1959). In addition, records of parasitism in the common crow, reported as a part of general surveys of bird parasites, are included in publications by Ransom (1909), Mayhew (1925), Cram (1927), Canavan (1929), Rankin (1946), Denton and Byrd (1951), Mawson (1956; 1957), Robinson (1954; 1955). This paper contains the results of a two-year study made in Ohio, during which 339 crows were examined for internal and external parasites. MATERIALS AND METHODS Juvenile and adult crows were shot in the field and wrapped individually in paper bags prior to transportation to the laboratory. -

University of Oklahoma
UNIVERSITY OF OKLAHOMA GRADUATE COLLEGE MACRONUTRIENTS SHAPE MICROBIAL COMMUNITIES, GENE EXPRESSION AND PROTEIN EVOLUTION A DISSERTATION SUBMITTED TO THE GRADUATE FACULTY in partial fulfillment of the requirements for the Degree of DOCTOR OF PHILOSOPHY By JOSHUA THOMAS COOPER Norman, Oklahoma 2017 MACRONUTRIENTS SHAPE MICROBIAL COMMUNITIES, GENE EXPRESSION AND PROTEIN EVOLUTION A DISSERTATION APPROVED FOR THE DEPARTMENT OF MICROBIOLOGY AND PLANT BIOLOGY BY ______________________________ Dr. Boris Wawrik, Chair ______________________________ Dr. J. Phil Gibson ______________________________ Dr. Anne K. Dunn ______________________________ Dr. John Paul Masly ______________________________ Dr. K. David Hambright ii © Copyright by JOSHUA THOMAS COOPER 2017 All Rights Reserved. iii Acknowledgments I would like to thank my two advisors Dr. Boris Wawrik and Dr. J. Phil Gibson for helping me become a better scientist and better educator. I would also like to thank my committee members Dr. Anne K. Dunn, Dr. K. David Hambright, and Dr. J.P. Masly for providing valuable inputs that lead me to carefully consider my research questions. I would also like to thank Dr. J.P. Masly for the opportunity to coauthor a book chapter on the speciation of diatoms. It is still such a privilege that you believed in me and my crazy diatom ideas to form a concise chapter in addition to learn your style of writing has been a benefit to my professional development. I’m also thankful for my first undergraduate research mentor, Dr. Miriam Steinitz-Kannan, now retired from Northern Kentucky University, who was the first to show the amazing wonders of pond scum. Who knew that studying diatoms and algae as an undergraduate would lead me all the way to a Ph.D. -

Isospora’ Lieberkuehni Labbe, 1894 (Apicomplexa: Sarcocystidae) and Its Taxonomic Implications
International Journal of Systematic and Evolutionary Microbiology (2001), 51, 767–772 Printed in Great Britain Phylogenetic position of a renal coccidium of the European green! frogs, ‘Isospora’ lieberkuehni Labbe, 1894 (Apicomplexa: Sarcocystidae) and its taxonomic implications 1 Department of David Modry! ,1,2 Jan R. S) lapeta,1,2 Milan Jirku/ ,2,3 Miroslav Obornı!k,2,3 Parasitology, University 2,3 1,2 of Veterinary and Julius Lukes) and Br) etislav Koudela Pharmaceutical Sciences, Palacke! ho 1-3, 612 42 Brno, Czech Republic Author for correspondence: David Modry! . Tel: j420 5 41562979. Fax: j420 5 748841. e-mail: modryd!vfu.cz 2,3 Institute of Parasitology, Czech Academy of Sciences2 and and Faculty of Biology, ‘Isospora’ lieberkuehni, an unusual isosporoid renal coccidium that parasitizes University of the European water frog was isolated from the edible frog, Rana kl. esculenta, South Bohemia3 , in the Czech Republic. Sequencing of the small-subunit (SSU) rRNA gene C) eske! Bude) jovice, Czech Republic showed that it belongs to the family Sarcocystidae, being closely related to a clade comprising members of the subfamily Toxoplasmatinae. The position within Sarcocystidae correlates with the mode of excystation via collapsible plates as postulated by previous authors. Phylogenetic, morphological and biological differences between ‘Isospora’ lieberkuehni and the other Stieda- body-lacking members of the genus Isospora justify separation of this ! coccidium on a generic level. Hyaloklossia Labbe, 1896 is the oldest available synonym -

The Intestinal Protozoa
The Intestinal Protozoa A. Introduction 1. The Phylum Protozoa is classified into four major subdivisions according to the methods of locomotion and reproduction. a. The amoebae (Superclass Sarcodina, Class Rhizopodea move by means of pseudopodia and reproduce exclusively by asexual binary division. b. The flagellates (Superclass Mastigophora, Class Zoomasitgophorea) typically move by long, whiplike flagella and reproduce by binary fission. c. The ciliates (Subphylum Ciliophora, Class Ciliata) are propelled by rows of cilia that beat with a synchronized wavelike motion. d. The sporozoans (Subphylum Sporozoa) lack specialized organelles of motility but have a unique type of life cycle, alternating between sexual and asexual reproductive cycles (alternation of generations). e. Number of species - there are about 45,000 protozoan species; around 8000 are parasitic, and around 25 species are important to humans. 2. Diagnosis - must learn to differentiate between the harmless and the medically important. This is most often based upon the morphology of respective organisms. 3. Transmission - mostly person-to-person, via fecal-oral route; fecally contaminated food or water important (organisms remain viable for around 30 days in cool moist environment with few bacteria; other means of transmission include sexual, insects, animals (zoonoses). B. Structures 1. trophozoite - the motile vegetative stage; multiplies via binary fission; colonizes host. 2. cyst - the inactive, non-motile, infective stage; survives the environment due to the presence of a cyst wall. 3. nuclear structure - important in the identification of organisms and species differentiation. 4. diagnostic features a. size - helpful in identifying organisms; must have calibrated objectives on the microscope in order to measure accurately. -

Control of Intestinal Protozoa in Dogs and Cats
Control of Intestinal Protozoa 6 in Dogs and Cats ESCCAP Guideline 06 Second Edition – February 2018 1 ESCCAP Malvern Hills Science Park, Geraldine Road, Malvern, Worcestershire, WR14 3SZ, United Kingdom First Edition Published by ESCCAP in August 2011 Second Edition Published in February 2018 © ESCCAP 2018 All rights reserved This publication is made available subject to the condition that any redistribution or reproduction of part or all of the contents in any form or by any means, electronic, mechanical, photocopying, recording, or otherwise is with the prior written permission of ESCCAP. This publication may only be distributed in the covers in which it is first published unless with the prior written permission of ESCCAP. A catalogue record for this publication is available from the British Library. ISBN: 978-1-907259-53-1 2 TABLE OF CONTENTS INTRODUCTION 4 1: CONSIDERATION OF PET HEALTH AND LIFESTYLE FACTORS 5 2: LIFELONG CONTROL OF MAJOR INTESTINAL PROTOZOA 6 2.1 Giardia duodenalis 6 2.2 Feline Tritrichomonas foetus (syn. T. blagburni) 8 2.3 Cystoisospora (syn. Isospora) spp. 9 2.4 Cryptosporidium spp. 11 2.5 Toxoplasma gondii 12 2.6 Neospora caninum 14 2.7 Hammondia spp. 16 2.8 Sarcocystis spp. 17 3: ENVIRONMENTAL CONTROL OF PARASITE TRANSMISSION 18 4: OWNER CONSIDERATIONS IN PREVENTING ZOONOTIC DISEASES 19 5: STAFF, PET OWNER AND COMMUNITY EDUCATION 19 APPENDIX 1 – BACKGROUND 20 APPENDIX 2 – GLOSSARY 21 FIGURES Figure 1: Toxoplasma gondii life cycle 12 Figure 2: Neospora caninum life cycle 14 TABLES Table 1: Characteristics of apicomplexan oocysts found in the faeces of dogs and cats 10 Control of Intestinal Protozoa 6 in Dogs and Cats ESCCAP Guideline 06 Second Edition – February 2018 3 INTRODUCTION A wide range of intestinal protozoa commonly infect dogs and cats throughout Europe; with a few exceptions there seem to be no limitations in geographical distribution. -

Enteric Protozoa in the Developed World: a Public Health Perspective
Enteric Protozoa in the Developed World: a Public Health Perspective Stephanie M. Fletcher,a Damien Stark,b,c John Harkness,b,c and John Ellisa,b The ithree Institute, University of Technology Sydney, Sydney, NSW, Australiaa; School of Medical and Molecular Biosciences, University of Technology Sydney, Sydney, NSW, Australiab; and St. Vincent’s Hospital, Sydney, Division of Microbiology, SydPath, Darlinghurst, NSW, Australiac INTRODUCTION ............................................................................................................................................420 Distribution in Developed Countries .....................................................................................................................421 EPIDEMIOLOGY, DIAGNOSIS, AND TREATMENT ..........................................................................................................421 Cryptosporidium Species..................................................................................................................................421 Dientamoeba fragilis ......................................................................................................................................427 Entamoeba Species.......................................................................................................................................427 Giardia intestinalis.........................................................................................................................................429 Cyclospora cayetanensis...................................................................................................................................430 -

Balantidium Coli
Intestinal Ciliates, Coccidia (Sporozoa) and Microsporidia Balantidium coli (Ciliates) Cyclospora cayetanensis (Coccidia, Sporozoa) Cryptosporidium parvum (Coccidia, Sporozoa) Enterocytozoon bieneusi (Microsporidia) Isospora belli (Coccidia, Sporozoa) Encephalitozoon intestinalis (Microsporidia) Balantidium coli Introduction: Balantidium coli is the only member of the ciliate (Ciliophora) family to cause human disease (Balantadiasis). It is the largest known protozoan causing human infection. Morphology: Trophozoites: Trophozoites are oval in shape and measure approximately 30-200 um. The main morphological characteristics are cilia, 2 types of nuclei; 1 macro (Kidney shaped) and 1 micronucleus and a large contractile vacuole. A funnel shaped cytosome can be seen near the anterior end. Cysts: The cyst is spherical or round ellipsoid and measures from 30-70 um. It contains 1 macro and 1 micronucleus. The cilia are present in young cysts and may be seen slowly rotating, but after prolonged encystment, the cilia disappear. Life Cycle: Definitive hosts are humans including a main reservoir are the pigs (It is estimated that 63-90 % harbor B. coli), rodents and other animals with no intermidiate hosts or vectors. Cysts (Infective stage) are responsible for transmission of balantidiasis through ingestion of contaminated food or water through the oral-fecal route. Following ingestion, excystation occurs in the small intestine, and the trophozoites colonize the large intestine. The trophozoites reside in the lumen of the large intestine of humans and animals, where they replicate by binary fission. Some trophozoites invade the wall of the colon and Parasitology Chapter 4 The Ciliates and Coccidia Page 1 of 9 multiply. Trophozoites undergo encystation to produce infective cysts. Mature cysts are passed with feces. -

Slide 1 This Lecture Is the Second Part of the Protozoal Parasites. in This LECTURE We Will Talk About the Apicomplexans SPECIFICALLY the Coccidians
Slide 1 This lecture is the second part of the protozoal parasites. In this LECTURE we will talk about the Apicomplexans SPECIFICALLY THE Coccidians. In the next lecture we will Lecture 8: Emerging Parasitic Protozoa part 1 (Apicomplexans-1: talk about the Plasmodia and Babesia Coccidia) Presented by Sharad Malavade, MD, MPH Original Slides by Matt Tucker, PhD HSC4933 1 Emerging Infectious Diseases Slide 2 These are the readings for this week. Readings-Protozoa pt. 2 (Coccidia) • Ch.8 (p. 183 [table 8.3]) • Ch. 11 (p. 301, 304-305) 2 Slide 3 Monsters Inside Me • Cryptosporidiosis (Cryptosporidum spp., Coccidian/Apicomplexan): Background: http://www.cdc.gov/parasites/crypto/ Video: http://animal.discovery.com/videos/monsters-inside-me- cryptosporidium-outbreak.html http://animal.discovery.com/videos/monsters-inside-me-the- cryptosporidium-parasite.html Toxoplasmosis (Toxoplasma gondii, Coccidian/Apicomplexan) Background: http://www.cdc.gov/parasites/toxoplasmosis/ Video: http://animal.discovery.com/videos/monsters-inside-me- toxoplasma-parasite.html 3 Slide 4 Learning objectives: Apicomplexan These are the learning objectives for this lecture. coccidia • Define basic attributes of Apicomplexans- unique characteristics? • Know basic life cycle and developmental stages of coccidian parasites • Required hosts – Transmission strategy – Infective and diagnostic stages – Unique character of reproduction • Know the common characteristics of each parasite – Be able to contrast and compare • Define diseases, high-risk groups • Determine diagnostic methods, treatment • Know important parasite survival strategies • Be familiar with outbreaks caused by coccidians and the conditions involved 4 Slide 5 This figure from the last lecture is just to show you the Taxonomic Review apicoplexans. This lecture we talk about the Coccidians. -

Canine Neosporosis: Clinical and Pathological ®Ndings and ®Rst Isolation of Neospora Caninum in Germany
Parasitol Res (2000) 86: 1±7 Ó Springer-Verlag 2000 ORIGINAL PAPER Martin Peters á Frank Wagner á Gereon Schares Canine neosporosis: clinical and pathological ®ndings and ®rst isolation of Neospora caninum in Germany Received: 8 August 1999 / Accepted: 30 August 1999 Abstract Neosporosis was diagnosed in an 11-week-old species. In cattle, N. caninum has been found to be as- puppy of the breed Kleiner MuÈ nsterlaÈ nder with sociated with abortions. Clinical neosporosis in dogs is progressive hindlimb paresis. Pathohistological and most commonly observed in congenitally infected pup- immunohistological examinations revealed a dissemi- pies in the ®rst 6 months of life (Ruehlmann et al. 1995). nated infection with Neospora caninum. Parasitic stages Characteristically these animals show a progressively were demonstrated in the brain, spinal cord, retina, ascending paresis, usually starting at the pelvic limbs, muscles, thymus, heart, liver, kidney, stomach, adrenal caused by severe polymyositis, polyradiculitis, and dis- gland, and skin. Immunohistochemistry investigations seminated meningoencephalomyelitis. Generalized in- were carried out using polyclonal rabbit antisera devel- fection with multiple organ involvement in puppies has oped against N. caninum tachyzoites and the recombi- been reported (Dubey et al. 1988a; Barber et al. 1996). nant bradyzoite-speci®c antigen BAG-5 of Toxoplasma Some puppies suered and eventually died of myo- gondii, which is known to cross-react with N. caninum carditis caused by the parasite (Dubey et al. 1988a; Odin bradyzoites. BAG-5 antibodies recognized tissue cysts and Dubey 1993; Barber et al. 1996; WeissenboÈ ck et al. within the CNS and some protozoan stages that were 1997). Besides multifocal CNS involvement and not surrounded by a visible cyst wall. -
Accelerated Reader List
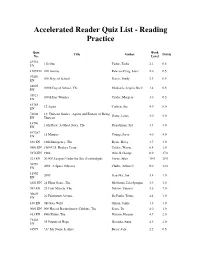
Accelerated Reader Quiz List - Reading Practice Quiz Book Title Author Points No. Level 43532 1 Is One Tudor, Tasha 2.1 0.5 EN 1209 EN 100 Acorns Palazzo-Craig, Janet 2.0 0.5 57450 100 Days of School Harris, Trudy 2.3 0.5 EN 41025 100th Day of School, The Medearis, Angela Shelf 1.4 0.5 EN 35821 100th Day Worries Cuyler, Margery 3.0 0.5 EN 61265 12 Again Corbett, Sue 4.9 8.0 EN 74604 13: Thirteen Stories...Agony and Ecstasy of Being Howe, James 5.0 9.0 EN Thirteen 14796 13th Floor: A Ghost Story, The Fleischman, Sid 4.4 4.0 EN 107287 15 Minutes Young, Steve 4.0 4.0 EN 661 EN 18th Emergency, The Byars, Betsy 4.7 4.0 9801 EN 1980 U.S. Hockey Team Coffey, Wayne 6.4 1.0 5976 EN 1984 Orwell, George 8.9 17.0 523 EN 20,000 Leagues Under the Sea (Unabridged) Verne, Jules 10.0 28.0 34791 2001: A Space Odyssey Clarke, Arthur C. 9.0 12.0 EN 11592 2095 Scieszka, Jon 3.8 1.0 EN 6651 EN 24-Hour Genie, The McGinnis, Lila Sprague 3.3 1.0 593 EN 25 Cent Miracle, The Nelson, Theresa 5.6 7.0 30629 26 Fairmount Avenue De Paola, Tomie 4.4 1.0 EN 166 EN 4B Goes Wild Gilson, Jamie 4.6 4.0 9001 EN 500 Hats of Bartholomew Cubbins, The Seuss, Dr. 4.0 1.0 413 EN 89th Kitten, The Nilsson, Eleanor 4.7 2.0 71428 95 Pounds of Hope Gavalda, Anna 4.3 2.0 EN 68579 "A" My Name Is Alice Bayer, Jane 2.2 0.5 Accelerated Reader Quiz List - Reading Practice Quiz Book Title Author Points No. -

"Plasmodium". In: Encyclopedia of Life Sciences (ELS)
Plasmodium Advanced article Lawrence H Bannister, King’s College London, London, UK Article Contents . Introduction and Description of Plasmodium Irwin W Sherman, University of California, Riverside, California, USA . Plasmodium Hosts Based in part on the previous version of this Encyclopedia of Life Sciences . Life Cycle (ELS) article, Plasmodium by Irwin W Sherman. Asexual Blood Stages . Intracellular Asexual Blood Parasite Stages . Sexual Stages . Mosquito Asexual Stages . Pre-erythrocytic Stages . Metabolism . The Plasmodium Genome . Motility . Recent History of Plasmodium Research . Evolution of Plasmodium . Conclusion Online posting date: 15th December 2009 Plasmodium is a genus of parasitic protozoa which infect Introduction and Description of erythrocytes of vertebrates and cause malaria. Their life cycle alternates between mosquito and vertebrate hosts. Plasmodium Parasites enter the bloodstream after a mosquito bite, Parasites of the genus Plasmodium are protozoans which and multiply sequentially within liver cells and erythro- invade and multiply within erythrocytes of vertebrates, and cytes before becoming male or female sexual forms. When are transmitted by mosquitoes. The motile invasive stages ingested by a mosquito, these fuse, then the parasite (merozoite, ookinete and sporozoite) are elongate, uni- multiplies again to form more invasive stages which are nucleate cells able to enter cells or pass through tissues, transmitted back in the insect’s saliva to a vertebrate. All using specialized secretory and locomotory organelles.