Protoplast Fusion of Citrus for Rootstock and Scion Improvement with Emphasis on Wide Hybridization
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Observations on Impietratura Disease Symptoms in Four Citrus Species
Observations on Impietratura Disease Symptoms in Four Citrus Species A. Caruso, M. Davino and G. Terranova ABSTRACT. Citrus impietratura disease affects citrus cultivars in the Mediterranean Basin. The indexing of impietratura disease is based on symptoms on indicator plants such as Volkamer lemon or grapefruit or on the inspection of the trees in the field. This requires many years. In this paper the effects of a severe isolate of impietratura on sweet orange, Volkamer lemon, rough lemon and Marsh Seedless grapefruit is reported. Our observations indicate that rough lemon is a better indicator of fruit symptoms than Volkamer lemon, grapefruit and sweet orange. Index words. biological assay, albedo gumming, rough lemon. In Italy cristacortis, concave gum servation for at least three years. Dif- and impietratura are widespread in ficulties in timely indexing of old line commercial orchards. The causal Navelina trees planted in Italy be- agents of these diseases have not been tween 1970 and 1985 resulted in the characterized and their detection is high percentage of infection (>30%) by still by means of biological assays. impietratura, concave gum and Due of its excellent bioagronomic psorosis A (4,6,13). and marketing character, in Italy the Rough lemon was initially consid- cultivation of the Navelina sweet ered to be tolerant to impietratura dis- orange spread rapidly in many citrus- ease (9), but later studies showed it to growing areas by growers, despite the be susceptible (7,8,12) and Catara and fact that the first trees imported pre- Scaramuzzi (5) suggested its use as an sented very mild flecking symptoms on alternative indicator. -

Rangpur Lime X Troyer Citrange, a Hybrid Citrus Rootstock for Closely Spaced Trees
Proc. Fla. State Hort. Soc. 99:33-35. 1986. RANGPUR LIME X TROYER CITRANGE, A HYBRID CITRUS ROOTSTOCK FOR CLOSELY SPACED TREES W. S. Castle A combination of diseases, repeated freezes, and other University of Florida, IFAS factors has reemphasized the importance of rootstocks in Citrus Research and Education Center Florida. Moreover, the effects of these factors illustrate the 700 Experiment Station Road inherent weaknesses in virtually all citrus rootstocks and Lake Alfred, FL 33850 the need to continually search for new, improved ones. Another recent trend related to rootstocks has been C. O. YOUTSEY the shift toward more closely spaced trees, particularly FDACS, Division of Plant Industry within the row (7,8). Rootstocks well-suited for dense plan Citrus Budwood Registration Bureau tings have not been available although such stocks are 3027 Lake Alfred Road being evaluated and one appears particularly promising Winter Haven, FL 33881 (1, 3, 4, 8). It is a hybrid of Rangpur lime and Troyer D. J. Hutchison citrange (RxT) and has been under study in Florida for 18 United States Department of Agriculture yr. During this period, trees on RxT have demonstrated Agricultural Research Service sufficient commercial potential to justify our presentation 2120 Camden Road in this report of their performance and a description of Orlando, FL 32803 RxT and its characteristics. Additional index words. Blight, tristeza, tree size control. History Dr. J. R. Furr, formerly a plant breeder with the U.S. Abstract. A hybrid of Rangpur lime (Citrus limonia Osb.) and Department of Agriculture (USDA) at Indio, California, Troyer citrange [ C. sinensis (L) Osb. -

Known Host Plants of Huanglongbing (HLB) and Asian Citrus Psyllid
Known Host Plants of Huanglongbing (HLB) and Asian Citrus Psyllid Diaphorina Liberibacter citri Plant Name asiaticus Citrus Huanglongbing Psyllid Aegle marmelos (L.) Corr. Serr.: bael, Bengal quince, golden apple, bela, milva X Aeglopsis chevalieri Swingle: Chevalier’s aeglopsis X X Afraegle gabonensis (Swingle) Engl.: Gabon powder-flask X Afraegle paniculata (Schum.) Engl.: Nigerian powder- flask X Atalantia missionis (Wall. ex Wight) Oliv.: see Pamburus missionis X X Atalantia monophylla (L.) Corr.: Indian atalantia X Balsamocitrus dawei Stapf: Uganda powder- flask X X Burkillanthus malaccensis (Ridl.) Swingle: Malay ghost-lime X Calodendrum capense Thunb.: Cape chestnut X × Citroncirus webberi J. Ingram & H. E. Moore: citrange X Citropsis gilletiana Swingle & M. Kellerman: Gillet’s cherry-orange X Citropsis schweinfurthii (Engl.) Swingle & Kellerm.: African cherry- orange X Citrus amblycarpa (Hassk.) Ochse: djerook leemo, djeruk-limau X Citrus aurantiifolia (Christm.) Swingle: lime, Key lime, Persian lime, lima, limón agrio, limón ceutí, lima mejicana, limero X X Citrus aurantium L.: sour orange, Seville orange, bigarde, marmalade orange, naranja agria, naranja amarga X Citrus depressa Hayata: shiikuwasha, shekwasha, sequasse X Citrus grandis (L.) Osbeck: see Citrus maxima X Citrus hassaku hort. ex Tanaka: hassaku orange X Citrus hystrix DC.: Mauritius papeda, Kaffir lime X X Citrus ichangensis Swingle: Ichang papeda X Citrus jambhiri Lushington: rough lemon, jambhiri-orange, limón rugoso, rugoso X X Citrus junos Sieb. ex Tanaka: xiang -

Eight New Somatic Hybrid Citrus Rootstocks with Potential For
HORTSCIENCE 27(9):1033-1036. 1992. pest resistance into horticulturally desirable rootstocks adapted to the various environ- mental conditions that exist in Florida. Eight New Somatic Hybrid Citrus Rough lemon, Volkamer lemon, and Rangpur have been used as rootstocks, be- Rootstocks with Potential for cause trees on these stocks are vigorous, high- yielding, and tristeza- and drought-tolerant Improved Disease Resistance (Castle, 1987; Castle et al., 1989). How- ever, their use has declined because they are Eliezer S. Louzada1, Jude W. Grosseti, Frederick G. Gmitter, Jr.2, highly susceptible to citrus blight. Cleopatra Beatriz Nielsen3, and J.L. Chandler3 mandarin has become an increasingly im- portant rootstock that is less susceptible to Department of Horticultural Sciences, Citrus Research and Education citrus blight than the rootstocks named above, Center, Institute of Food and Agricultural Sciences, University of tolerant to tristeza, and cold-hardy. Trees Florida, 700 Experiment Station Road, Lake Alfred, FL 33850 grafted on Cleopatra mandarin are moder- ately vigorous but are slow to reach full 2 Xiu Xin Deng bearing potential (Castle et al., 1989). Sweet Department of Horticulture, Huazhong Agricultural University, Wuhan orange is tolerant of citrus blight, but is not Hubei 430070, People’s Republic of China used as a rootstock due to Phytophthora sus- ceptibility. Somatic hybridization of Cleo- Nicasio Tusa4 patra mandarin with rough lemon, Volkamer lemon, and Rangpur; ‘Hamlin’ sweet orange Centro di Studio per il Miglioramento Genetico degli Agrumi, Consiglio with Rangpur; and ‘Valencia’ sweet orange Nazionale delle Ricerche, Male delle Scienze, Palenno 90128, Sicily with Carrizo citrange was attempted to gen- Additional index words. -

119-123 (2008) ว. วิทย. กษ. 39 : 3 (พเศษิ ) : 119-123 (2551)
Agricultural Sci. J. 39 : 3 (Suppl.) : 119-123 (2008) ว. วิทย. กษ. 39 : 3 (พเศษิ ) : 119-123 (2551) การยับยั้งเชื้อราสาเหตุโรคพืชดวยสารสกัดจากพืชในสกุล Rutaceae 16 ชนิด Antimicrobial activity of 16 plant extracts of the Rutaceae family against phytopathogenic fungi เนตรนภสิ เขียวขาํ 1, Harald Greger2 และ สมศิริ แสงโชต1ิ Netnapis Khewkhom1, Harald Greger2 and Somsiri Shangchote1 Abstract The lipophilic extract of 16 plant extracts of the Rutaceae family was selected for investigation of antifungal properties. Antifungal active compounds bioautography bioassays against Cladosporium herbarum have been detected. Comparative studies of Toddalia sp. (leaves), Limonia acidissima (leaves), Vepris bilocularis (leaves), Coleonema pulchellum (roots), Triphasia trifoliata (leaves), Pleiospermum alatum (leaves and stem), Acronychia pedunculata (leaves), and Atalantia sp. (leaves) extract showed clear inhibition zones on TLC plates against C. herbarum. In microdilution bioassay, the leaf extracts of Fortunella hindsii exhibited the strongest fungicidal activity with a MIC value at 312.5 μg/mL and showed an inhibition of spore germination at EC50 of 114 μg/mL for Botrytis cinerea. L. acidissima leaf extracts displayed a MIC of 1250 μg/mL and showed an inhibition of spore germination with EC50 0 values at 589 μg/mL for B. cinerea. The leaf extracts of two different collections of Glycosmis mauritiana (RUT 213/7) and (RUT 400) revealed clear differences for the antifungal activity: the EC50 value of the former was at 70 μg/mL, but 1249 μg/mL for the latter. G. mauritiana (RUT 213/7) revealed clear differences for the antifungal activity: the EC50 value at 70 μg/mL. Stem extracts of Pleiospermum alatum showed activities with values at EC50 262 μg/mL against B. -
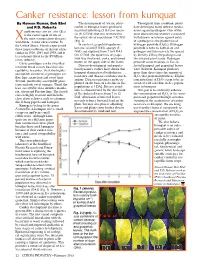
Canker Resistance: Lesson from Kumquat by Naveen Kumar, Bob Ebel the Development of Asiatic Citrus Throughout Their Evolution, Plants and P.D
Canker resistance: lesson from kumquat By Naveen Kumar, Bob Ebel The development of Asiatic citrus Throughout their evolution, plants and P.D. Roberts canker in kumquat leaves produced have developed many defense mecha- anthomonas citri pv. citri (Xcc) localized yellowing (5 DAI) or necro- nisms against pathogens. One of the is the causal agent of one of sis (9-12 DAI) that was restricted to most characteristic features associated the most serious citrus diseases the actual site of inoculation 7-12 DAI with disease resistance against entry X (Fig. 2). of a pathogen is the production of worldwide, Asiatic citrus canker. In the United States, Florida experienced In contrast, grapefruit epidermis hydrogen peroxide (H2O2). Hydrogen three major outbreaks of Asiatic citrus became raised (5 DAI), spongy (5 peroxide is toxic to both plant and canker in 1910, 1984 and 1995, and it DAI) and ruptured from 7 to 8 DAI. pathogen and thus restricts the spread is a constant threat to the $9 billion On 12 DAI, the epidermis of grape- by directly killing the pathogen and citrus industry. fruit was thickened, corky, and turned the infected plant tissue. Hydrogen Citrus genotypes can be classified brown on the upper side of the leaves. peroxide concentrations in Xcc-in- into four broad classes based on sus- Disease development and popula- fected kumquat and grapefruit leaves ceptibility to canker. First, the highly- tion dynamics studies have shown that were different. Kumquat produces susceptible commercial genotypes are kumquat demonstrated both disease more than three times the amount of Key lime, grapefruit and sweet lime. -

PRUNING GUIDELINES Tools: Loppers, Saw, Clippers 10% Bleach and Water Solution
Varieties - choose one that you will want to eat often, as you will have them much of the year (Four Winds Citrus Variety Chart link in your Resources list) - certain citrus mature earlier than others (see early ripening handout) - Unique varieties: - Blood orange: red flesh is antioxidant rich. Often sweeter than other oranges - Yuzu: very little, but very flavorful juice used by chefs. Believed to be a cross between a sour mandarin and ichang papeda - Keiffir lime: regular lime with bumpy skin. Attractive tree with segmented leaves that are extremely fragrant and prized by chefs. - Buddhas hand: not much juice, but very fragrant pith and rind. Odd shaped and often used as an ornamental - Australian finger lime: oblong green lime with many small lime-flavored orbs inside. Called "citrus caviar" Standard varieties of citrus trees often grow to a height of 20 to 30 feet and the canopy -- or width of a tree -- can spread to 18 to 30 feet depending on the variety. Dwarf citrus trees are significantly shorter and narrower, which provides greater flexibility in planting location. Most varieties top out at 8 feet in height with a proportionally smaller canopy. Despite the differences in height and width, regular, semi-dwarf and dwarf citrus varieties produce the same size fruit. -First off DO NOT PRUNE! Those damaged leaves can actually provide protection for the plant until the air warms up. The plant needs to rally and recover. Pruning might just put it over the edge. Sometimes the plants must remain with that ‘raggedy’ appearance until as late as June and in some cases a full year. -

The Remote Citroid Fruit Tree Glycosmis Pentaphylla Is a Host of Citrus Leprosis Virus C and Exhibits Novel Leprosis Symptoms Fi
031_TESTI_DN_499 25-06-2009 12:55 Pagina 499 Journal of Plant Pathology (2009), 91 (2), 499-505 Edizioni ETS Pisa, 2009 499 DISEASE NOTE DISEASE NOTE THE REMOTE CITROID FRUIT TREE FIRST REPORT OF PHOMOPSIS GLYCOSMIS PENTAPHYLLA IS A HOST ACTINIDIAE CAUSING CANKERS ON OF CITRUS LEPROSIS VIRUS C AND SHOOTS OF KIWIFRUIT IN GREECE EXHIBITS NOVEL LEPROSIS SYMPTOMS T. Thomidis J. Freitas-Astúa1,2, A.L. Fadel2, R. Antonioli-Luizon2, M. Bas- tianel2, V.M. Novelli2, E.W. Kitajima3 and M.A. Machado2 Alexander Technological Education Institute of Thessaloniki, School of Agricultural Technology, Department 1Embrapa Cassava and Tropical Fruits, s/n, Cruz das Almas, BA, of Crop Production, 57400 Sindos, Thessaloniki, Greece Brazil, 44380-000 2Centro APTA Citros Sylvio Moreira-IAC, CP 4, Cordeirópolis, SP, In June and through summer 2007, wilted and blighted Brazil,13490-970 shoots with distinct dark cankers were observed on ki- 3Departamento de Entomologia, Fitopatologia e Zoologia Agrícola, wifruit plants of cvs Hayward and Tsechelidis in the ESALQ, CP9, Piracicaba, SP, Brazil, 13418-900 province of Imathia (Greece). Isolations made on acidified potato dextrose agar yielded a fungus which was identified Citrus leprosis virus C (CiLV-C), the causal agent of as Phomopsis actinidiae (Henn.) Died. on the basis of mor- one of the main virus disease of citrus in Brazil, is consid- phological and sporulation characters. Circular chalk ered to have a narrow host range. However, recent studies white-colored aerial mycelial mats developed in culture af- ter incubation at 25°C for 7 days. After additional 3-4 have shown that some weeds, hedgerow and windbreak weeks, black, spherical or bluntly conical pycnidia 230-500 plants can host CiLV-C, thus may play a role in the epi- µm in size bearing a- and b-conidia were formed all over demiology of the disease (Bastianel et al., 2006). -

Citrus from Seed?
Which citrus fruits will come true to type Orogrande, Tomatera, Fina, Nour, Hernandina, Clementard.) from seed? Ellendale Tom McClendon writes in Hardy Citrus Encore for the South East: Fortune Fremont (50% monoembryonic) “Most common citrus such as oranges, Temple grapefruit, lemons and most mandarins Ugli Umatilla are polyembryonic and will come true to Wilking type. Because most citrus have this trait, Highly polyembryonic citrus types : will mostly hybridization can be very difficult to produce nucellar polyembryonic seeds that will grow true to type. achieve…. This unique characteristic Citrus × aurantiifolia Mexican lime (Key lime, West allows amateurs to grow citrus from seed, Indian lime) something you can’t do with, say, Citrus × insitorum (×Citroncirus webberii) Citranges, such as Rusk, Troyer etc. apples.” [12*] Citrus × jambhiri ‘Rough lemon’, ‘Rangpur’ lime, ‘Otaheite’ lime Monoembryonic (don’t come true) Citrus × limettioides Palestine lime (Indian sweet lime) Citrus × microcarpa ‘Calamondin’ Meyer Lemon Citrus × paradisi Grapefruit (Marsh, Star Ruby, Nagami Kumquat Redblush, Chironja, Smooth Flat Seville) Marumi Kumquat Citrus × sinensis Sweet oranges (Blonde, navel and Pummelos blood oranges) Temple Tangor Citrus amblycarpa 'Nasnaran' mandarin Clementine Mandarin Citrus depressa ‘Shekwasha’ mandarin Citrus karna ‘Karna’, ‘Khatta’ Poncirus Trifoliata Citrus kinokuni ‘Kishu mandarin’ Citrus lycopersicaeformis ‘Kokni’ or ‘Monkey mandarin’ Polyembryonic (come true) Citrus macrophylla ‘Alemow’ Most Oranges Citrus reshni ‘Cleopatra’ mandarin Changshou Kumquat Citrus sunki (Citrus reticulata var. austera) Sour mandarin Meiwa Kumquat (mostly polyembryonic) Citrus trifoliata (Poncirus trifoliata) Trifoliate orange Most Satsumas and Tangerines The following mandarin varieties are polyembryonic: Most Lemons Dancy Most Limes Emperor Grapefruits Empress Tangelos Fairchild Kinnow Highly monoembryonic citrus types: Mediterranean (Avana, Tardivo di Ciaculli) Will produce zygotic monoembryonic seeds that will not Naartje come true to type. -

Supplementary Material for RUSSELL, DYRANA N., JAWWAD A
Supplementary Material for RUSSELL, DYRANA N., JAWWAD A. QURESHI, SUSAN E. HALBERT AND PHILIP A. STANSLY−Host Suitability of Citrus and Zanthoxylum Spp. for Leuronota fagarae and Diaphorina citri (Hemiptera: Psylloidea). Florida Entomologist 97(4) (December 2014) at http://purl.fcla.edu/fcla/entomologist/browse Corresponding author: Dr. J. A. Qureshi University of Florida/IFAS Southwest Florida Research and Education Center (SWFREC) 2685 SR 29N, Immokalee, Fl 34142, USA Phone: (239) 658-3400 Fax: (239) 658-3469 E-mail: [email protected] ABSTRACT Leuronota fagarae Burckhardt (Hemiptera: Psylloidea), an exotic psyllid described from South America, was first observed in 2001on a citrus relative Zanthoxylum fagara (L.) Sarg. (Sapindales: Rutaceae) in southern Florida. Diaphorina citri Kuwayama (Hemiptera: Psylloidea) is principal vector of the bacteria ‘Candidatus Liberibacter spp.’ causal agent of huanglongbing (HLB) or citrus greening disease. Both vector and disease are now well established in Florida and also reported throughout the Americas and Asia. The host range of D. citri is limited to citrus and some rutaceous relatives. Additional vectors and host plants could accelerate spread of HLB in citrus and threaten endangered species such as Zanthoxylum coriaceum A. Rich. and Zanthoxylum flavum Vahl. Experiments were conducted to evaluate adult survival, reproduction and nymphal development of psyllids on 3 Citrus and 4 Zanthoxylum species as well as orange jasmine, Murraya paniculata (Syn. M. exotica) (Sapindales: Rutaceae), a common ornamental and preferred host of D. citri. Leuronota fagarae in single male−female pairs at 24 °C lived an average 4-47 days, 4-12 fold longer on Zanthoxylum spp. (except Z. flavum) than on citrus. -

Tetraploid Citrumelo 4475 Rootstocks Improve Diploid Common
www.nature.com/scientificreports OPEN Tetraploid Citrumelo 4475 rootstocks improve diploid common clementine tolerance to long‑term nutrient defciency Julie Oustric1*, Stéphane Herbette2, Yann Quilichini3, Raphaël Morillon4,5, Jean Giannettini1, Liliane Berti1 & Jérémie Santini1 Nutrient defciency alters growth and the production of high‑quality nutritious food. In Citrus crops, rootstock technologies have become a key tool for enhancing tolerance to abiotic stress. The use of doubled diploid rootstocks can improve adaptation to lower nutrient inputs. This study investigated leaf structure and ultrastructure and physiological and biochemical parameters of diploid common clementine scions (C) grafted on diploid (2x) and doubled diploid (4x) Carrizo citrange (C/CC2x and C/CC4x) and Citrumelo 4475 (C/CM2x and C/CM4x) rootstocks under optimal fertigation and after 7 months of nutrient defciency. Rootstock ploidy level had no impact on structure but induced changes in the number and/or size of cells and some cell components of 2x common clementine leaves under optimal nutrition. Rootstock ploidy level did not modify gas exchanges in Carrizo citrange but induced a reduction in the leaf net photosynthetic rate in Citrumelo 4475. By assessing foliar damage, changes in photosynthetic processes and malondialdehyde accumulation, we found that C/CM4x were less afected by nutrient defciency than the other scion/rootstock combinations. Their greater tolerance to nutrient defciency was probably due to the better performance of the enzyme‑based antioxidant system. Nutrient defciency had similar impacts on C/CC2x and C/CC4x. Tolerance to nutrient defciency can therefore be improved by rootstock polyploidy but remains dependent on the rootstock genotype. Fruit crops, especially citrus fruits, require large amounts of fertilizers to ensure good production and fruit qual- ity. -

The New York Botanical Garden
Vol. XV DECEMBER, 1914 No. 180 JOURNAL The New York Botanical Garden EDITOR ARLOW BURDETTE STOUT Director of the Laboratories CONTENTS PAGE Index to Volumes I-XV »33 PUBLISHED FOR THE GARDEN AT 41 NORTH QUBKN STRHBT, LANCASTER, PA. THI NEW ERA PRINTING COMPANY OFFICERS 1914 PRESIDENT—W. GILMAN THOMPSON „ „ _ i ANDREW CARNEGIE VICE PRESIDENTS J FRANCIS LYNDE STETSON TREASURER—JAMES A. SCRYMSER SECRETARY—N. L. BRITTON BOARD OF- MANAGERS 1. ELECTED MANAGERS Term expires January, 1915 N. L. BRITTON W. J. MATHESON ANDREW CARNEGIE W GILMAN THOMPSON LEWIS RUTHERFORD MORRIS Term expire January. 1916 THOMAS H. HUBBARD FRANCIS LYNDE STETSON GEORGE W. PERKINS MVLES TIERNEY LOUIS C. TIFFANY Term expire* January, 1917 EDWARD D. ADAMS JAMES A. SCRYMSER ROBERT W. DE FOREST HENRY W. DE FOREST J. P. MORGAN DANIEL GUGGENHEIM 2. EX-OFFICIO MANAGERS THE MAYOR OP THE CITY OF NEW YORK HON. JOHN PURROY MITCHEL THE PRESIDENT OP THE DEPARTMENT OP PUBLIC PARES HON. GEORGE CABOT WARD 3. SCIENTIFIC DIRECTORS PROF. H. H. RUSBY. Chairman EUGENE P. BICKNELL PROF. WILLIAM J. GIES DR. NICHOLAS MURRAY BUTLER PROF. R. A. HARPER THOMAS W. CHURCHILL PROF. JAMES F. KEMP PROF. FREDERIC S. LEE GARDEN STAFF DR. N. L. BRITTON, Director-in-Chief (Development, Administration) DR. W. A. MURRILL, Assistant Director (Administration) DR. JOHN K. SMALL, Head Curator of the Museums (Flowering Plants) DR. P. A. RYDBERG, Curator (Flowering Plants) DR. MARSHALL A. HOWE, Curator (Flowerless Plants) DR. FRED J. SEAVER, Curator (Flowerless Plants) ROBERT S. WILLIAMS, Administrative Assistant PERCY WILSON, Associate Curator DR. FRANCIS W. PENNELL, Associate Curator GEORGE V.