Population Structure and Growth Curve of Acanthopleura Gemmata (Mollusca: Polyplacophora) in the Northwestern Red Sea
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Download Preprint
1 Mobilising molluscan models and genomes in biology 2 Angus Davison1 and Maurine Neiman2 3 1. School of Life Sciences, University Park, University of Nottingham, NG7 2RD, UK 4 2. Department of Biology, University of Iowa, Iowa City, IA, USA and Department of Gender, 5 Women's, and Sexuality Studies, University of Iowa, Iowa, City, IA, USA 6 Abstract 7 Molluscs are amongst the most ancient, diverse, and important of all animal taxa. Even so, 8 no individual mollusc species has emerged as a broadly applied model system in biology. 9 We here make the case that both perceptual and methodological barriers have played a role 10 in the relative neglect of molluscs as research organisms. We then summarize the current 11 application and potential of molluscs and their genomes to address important questions in 12 animal biology, and the state of the field when it comes to the availability of resources such 13 as genome assemblies, cell lines, and other key elements necessary to mobilising the 14 development of molluscan model systems. We conclude by contending that a cohesive 15 research community that works together to elevate multiple molluscan systems to ‘model’ 16 status will create new opportunities in addressing basic and applied biological problems, 17 including general features of animal evolution. 18 Introduction 19 Molluscs are globally important as sources of food, calcium and pearls, and as vectors of 20 human disease. From an evolutionary perspective, molluscs are notable for their remarkable 21 diversity: originating over 500 million years ago, there are over 70,000 extant mollusc 22 species [1], with molluscs present in virtually every ecosystem. -

<I>Acanthopleura Gemmata</I>
NOTES 339 Mauri, M. and E. Orlando. 1983. Variability of zinc and manganese concentrations in relation to sex and season in the bivalve Donax trunculus. Mar. Pollut. Bull. 14: 342-346. McConchie, D. and L. M. Lawrance. 1991. The origin of high cadmium loads in some bivalves molluscs from Shark Bay, Western Australia: a new mechanism for cadmium uptake by filter feeding organisms. Arch. Environ. Contam. Toxicol. 21: 303-310. Nugegoda, D. and P. S. Rainbow. 1987. The effect of temperature on zinc regulation by the decapod crustacean Palaemon elegans Rathke. Ophelia 27: 17-30. Orlando, E. ]985. Valutazione dell'inquinamento marino da metalli pesanti tramite ]'uso di indicatori biologici. Oebalia XI: 93-100. Orren, M. J., G. A. Eagle, H. F.-K. O. Hennig and A. Green. 1980. Variations in trace metal content of the mussel Choromytilus meridionalis (Kr.) with season and sex. Mar. Pollut. Bull. 11: 253- 257. Rainbow, P. S. and A. G. Scott. 1979. Two heavy metal-binding proteins in the midgut gland of the crab Carcinus maenas. Mar. BioI. 55: 143-150. DATE ACCEPTED: April 13, ]994. ADDRESSES: (K.F.) Western Australian Marine Research Laboratories, PO Box 20, North Beach 6020, Australia; (CS.) Chemistry Centre of Western Australia, 125 Hay Street, East Perth 6004, Australia; (M.J.) Health Department of Western Australia, 189 Royal Street, East Perth 6004, Aus- tralia. BULLETINOF MARINESCIENCE,56( I): 339-343, 1995 KARYOLOGICAL STUDIES ON THE COMMON ROCKY EGYPTIAN CHITON, ACANTHOPLEURA GEMMATA (POLYPLACOPHORA: MOLLUSCA) Ahmed E. Yaseen, Abdel-Baset M. Ebaid and I. S. Kawashti Acanthopleura gemmata (Blainville, 1925) is one of the commonest polypla- cophoran species and is very common along the Egyptian coasts (northwestern part of the Red Sea) (Soliman and Habib, 1990). -

Polyplacophora: the Sesoko Gtation
The malacological society of Japan VEN US Jour. MHIac.} Hn (Jap, - Vol. 5S, Nu. 1 c1996}/ 41 49 -vs mazanog= u 7 ti l K 4 twoi)ts - itimaptL ptideptkA ・ ( pan-E11i!;( ftcma j(4}!: J,!UKJI(A7mpt}ttpt) of Distribution of Four Species Acanthopteura in Sesoko Island, Okinawa' (Polyplacophora:Chitonidae) Eiji YosHIoKA and Yasuhiro NAKAsHIMA (Kobe Yamate Women's College, Nakayamate-dori 6, Kobe 650 and Faculty of Science, Kyoto University, Kyoto 606-Ol) Abstract: Horizontal and vertical distribution of chitons, Acanthopleura gemmata. A. have been investigated. In the horizontal aspect, toochooana.A. tenuispinosa and A. miles, in the sheltered-exposed A. distributes they have specific distribution gradation. gemmata all rock shores around the island. A. Ioochooana distributes almost al1 around the island area. A. tenuispinosa mainly distributes the most sheltered area. except the most exposed In the vertical aspect, their A, miies mainly distributes the most exposed area. habitats significant except between are mostly overlapping, but their mean height have difference is lower, that of A. A. tenuispinosa and A. mites. The mean height of A, gemmata toochooana is middle, those of A, tenuispinosa and A, miles are higher in their habitat counts together, those species have specific zone. Puttingthe horizontal and vertical a ¢ four `space sharing' the dimensions although their habitat area and range overlap in in twe the most cases. Introduction Ferreira (1986) reviewed the genus Acanthopleura elaborately, and surnmarized the distri- bution of Acanthqpleura spp. According to his article, the distribution of three species, A. gemrnata (Fig. 1-A), A. Ioochooana (Fig. 1-B) and A. -
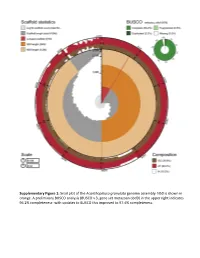
Snail Plot of the Acanthopleura Granulata Genome Assembly
Supplementary Figure 1: Snail plot of the Acanthopleura granulata genome assembly. N50 is shown in orange. A preliminary BUSCO analysis (BUSCO v.3, gene set metazoan obd9) in the upper right indicates 96.2% completeness- with updates to BUSCO this improved to 97.4% completeness. 1e+03 ● 1e+01 Mb N50), log(scaffold log(scaffold 1e−01 1e−03 20 40 60 80 100 BUSCO score, % Supplementary Figure 2: An illustration of the quality of available molluscan genomes as a scatter plot of BUSCO completeness score and a log transform of the N50 of scaffolds in megabases for each genome. The A. granulata genome is the orange circle, bivalves are purple triangles, gastropods are green squares, and cephalopods are blue stars. Grey diamonds are representative genomes of other lophotrochozoans (specifically those used as outgroups in the homolog gene searches in this study). 100 Acanthopleura granulata 93 Acanthopleura gemmata Rhyssoplax olivaceus 100 Chitonina 100 Chiton marmoratus Chitonida 100 Lepidozona mertensii 100 Katharina tunicata Mopalia muscosa Polyplacophora 100 100 Nuttallochiton mirandus Acanthochitonina 100 Cryptoplax larvaeformis 100 100 Acanthochitona crinita Callochiton sp. Leptochiton asellus Lepidopleurida 100 Hanleya nagelfar Prochaetoderma californicum 100 100 Chaetoderma nitidulum Caudofoveata 100 Falcidens sagittiferus Scutopus ventrolineatus 84 Epimenia babai 100 54 100 Apodomenia enigmatica 100 Neomenia megatrapezata Solenogastres 100 Stylomenia sulcodoryata 100 Amphimeniidae sp. 100 Octopus bimaculoides 100 Euprymna scolopes Cephalopoda -

Curaçao and Other
STUDIES ON THE FAUNA OF CURAÇAO AND OTHER CARIBBEAN ISLANDS: No. 137. Polyplacophora of the Caribbean Region by P. Kaas (Zoologisch Laboratorium, Utrecht) Contents Page Figure Plate INTRODUCTION 3 Materials 4 Historical review 5 Distribution (Tables 1 & 2) 8 SYSTEM ATICS 14 Lepidopleurida 15 Lepidopleuridae 15 Lepidopleurus Risso, 1826 16 1 pergranatus (Dall, 1889) 16 1-6 2 — binghami Boone, 1928 18 7-12 Hanleya Gray, 1857 20 3 — tropicalis (Dall, 1881) 20 13-18 Chitonida 21 Lepidochitonidae 22 Lepidochitona Gray, 1821 22 4 22 1 liozonis (Dall & Simpson, 1901) .... 19-40 I, 5 - rosea sp. n 27 41-49 Mopaliidae 29 Ceratozona Dall, 1882 29 6 rugosa (Sowerby, 1840) 29 50-54 1,2-3 Cryptoplacidae 33 Cryptoconchus Burrow, 1815 34 7 - floridanus (Dall, 1889) 34 55-57 I, 4-5 2 Page Figure Plate Acanthochitona Gray, 1821 37 8 hemphilli {Pilsbry, 1893) 38 58-64 II, 1-2 9 rhodea (Pilsbry, 1893) 42 65-71 bonairensis 44 72-73 10 n.sp III, 1-2 11 spiculosa (Reeve, 1847) 46 74-81 12 pygmaea (Pilsbry, 1893) 49 82-89 13 - 51 90-94 elongata n.sp 11,3 - 53 14 interfissa n.sp 95-107 Choneplax (Carpenter MS) Dall, 1882 . 55 15 lata (Guilding, 1829) 55 108-116 11,4 Ischnochitonidae 58 Calloplax Thiele, 1909 59 16 janeirensis (Gray, 1828) 60 117-123 IV, 1-2 Chaetopleura Shuttleworth, 1853 .... 63 17 apiculata (Say, 1830) 63 124-128 IV, 3-6 Ischnochiton Gray, 1847 66 B. 68 129-134 18 purpurascens (C. Adams, 1845) ... V, 1 19 - floridanus Pilsbry, 1892 72 135-136 V, 2 20 — boogii Haddon, 1886 74 137—150 21 - striolatus Gray, 1828 77 151-166 V, 3-4 - B. -

Mollusca: Polyplacophora)
Research Ideas and Outcomes 6: e60446 doi: 10.3897/rio.6.e60446 Project Report Large-scale project ‘Chiton of the Mexican Tropical Pacific’: Chiton articulatus (Mollusca: Polyplacophora) Omar Hernando Avila-Poveda ‡, § ‡ Facultad de Ciencias del Mar (FACIMAR), Universidad Autonoma de Sinaloa (UAS), Mazatlan, Sinaloa, Mexico § Direccion de Catedras-CONACYT, Consejo Nacional de Ciencia y Tecnologia (CONACYT), Ciudad de Mexico, Mexico Corresponding author: Omar Hernando Avila-Poveda ([email protected]) Reviewable v 1 Received: 06 Nov 2020 | Published: 10 Nov 2020 Citation: Avila-Poveda OH (2020) Large-scale project ‘Chiton of the Mexican Tropical Pacific’: Chiton articulatus (Mollusca: Polyplacophora). Research Ideas and Outcomes 6: e60446. https://doi.org/10.3897/rio.6.e60446 Abstract The marine mollusc, commonly called sea cockroach or chiton Chiton articulatus, is a mollusc belonging to the group known as Polyplacophora because its shell is composed of eight individual plates. This mollusc inhabits the rocky intertidal shore of the Mexican Tropical Pacific, where it is endemic. It has ecological, but also economic, importance. Ecologically, it is the preferred food of the snail Plicopurpura pansa, a protected species, in the cultural heritage of the country. Additionally, it is a basibiont (generates substrate for other individuals) that maintains the biodiversity of the Region. Economically, it has changed from artisanal consumption to become a culinary tourist attraction, offered at restaurants as an exotic and aphrodisiac dish, in tourist places like Huatulco or Acapulco. Despite being an exploited resource for decades, little is known about its life history. The Mexican Authorities have not yet recognised this mollusc as a fishing resource, so that it does not have any law that controls its extraction, sale and consumption, putting at risk the recruitment, survival and permanence of this species. -

Illustrated Summary of Chiton Terminology
©Zoologische Staatssammlung München/Verlag Friedrich Pfeil; download www.pfeil-verlag.de SPIXIANA 33 2 171–194 München, November 2010 ISSN 0341–8391 Illustrated summary of chiton terminology (Mollusca, Polyplacophora) Enrico Schwabe Schwabe, E. 2010. Illustrated summary of chiton terminology (Mollusca, Poly- placophora). Spixiana 33 (2): 171-194. The aims of the present paper are to summarize and offer a standard set of ter- minology used to describe morphological and partly anatomical (e. g. the radula) characters of Polyplacophora. To make the understanding of some previously misused terms easier, the identification and description of relevant parts of the animal are illustrated and discussed in context of additional literature. Enrico Schwabe, Bavarian State Collection of Zoology, Muenchhausenstr. 21, 81247 Munich, Germany; e-mail: [email protected] Introduction “An den Tentakeln des Kopfes befinden sich Riech- organe [On the tentacles of the head there are Chitons are a group of basal, exclusively marine olfactory organs.].” However, chitons do not have molluscs, which have not significantly changed cephalic tentacles at all, and certainly not olfactory their bauplan during more than 300 million years tentacles! of evolution. Their more or less solid, dorsal plates This short contribution intends to clarify the termi- have been preserved in numerous fossil records and nology of chitons and to present a detailed general allow researchers a direct comparison with living description of chiton morphology, to summarise for species, and accordingly there is a high number amateurs, students or scientists the present stage of described fossil and recent taxa. At present we of scientific knowledge of a fascinating group of count for about 930 recent species (see Schwabe animals. -

Foraging Tactics in Mollusca: a Review of the Feeding Behavior of Their Most Obscure Classes (Aplacophora, Polyplacophora, Monoplacophora, Scaphopoda and Cephalopoda)
Oecologia Australis 17(3): 358-373, Setembro 2013 http://dx.doi.org/10.4257/oeco.2013.1703.04 FORAGING TACTICS IN MOLLUSCA: A REVIEW OF THE FEEDING BEHAVIOR OF THEIR MOST OBSCURE CLASSES (APLACOPHORA, POLYPLACOPHORA, MONOPLACOPHORA, SCAPHOPODA AND CEPHALOPODA) Vanessa Fontoura-da-Silva¹, ², *, Renato Junqueira de Souza Dantas¹ and Carlos Henrique Soares Caetano¹ ¹Universidade Federal do Estado do Rio de Janeiro, Instituto de Biociências, Departamento de Zoologia, Laboratório de Zoologia de Invertebrados Marinhos, Av. Pasteur, 458, 309, Urca, Rio de Janeiro, RJ, Brasil, 22290-240. ²Programa de Pós Graduação em Ciência Biológicas (Biodiversidade Neotropical), Universidade Federal do Estado do Rio de Janeiro E-mails: [email protected], [email protected], [email protected] ABSTRACT Mollusca is regarded as the second most diverse phylum of invertebrate animals. It presents a wide range of geographic distribution patterns, feeding habits and life standards. Despite the impressive fossil record, its evolutionary history is still uncertain. Ancestors adopted a simple way of acquiring food, being called deposit-feeders. Amongst its current representatives, Gastropoda and Bivalvia are two most diversely distributed and scientifically well-known classes. The other classes are restricted to the marine environment and show other limitations that hamper possible researches and make them less frequent. The upcoming article aims at examining the feeding habits of the most obscure classes of Mollusca (Aplacophora, Polyplacophora, Monoplacophora, Scaphoda and Cephalopoda), based on an extense literary research in books, journals of malacology and digital data bases. The review will also discuss the gaps concerning the study of these classes and the perspectives for future analysis. -

Mollusca: Polyplacophora
Aquaculture and Fisheries Studies Research Open Volume 3 Issue 1 Short Report Chitons (Mollusca: Polyplacophora) from the Venezuelan Coasts, Southern Caribbean: A Checklist and Brief Review in Advance of Their Knowledge Ignacio Agudo-Padrón* Geographer & Research Malacologist, CEO Project “Avulsos Malacológicos – AM, Brazil *Corresponding author: Dr. Ignacio Agudo-Padrón, Geographer & Research Malacologist, CEO Project “Avulsos Malacológicos – AM”, Caixa Postal (P.O.Box) 010, 88010-970 Centro, Florianópolis, Santa Catarina/SC, Brazil; Email: [email protected] Received: December 31, 2020; Accepted: January 06, 2021; Published: January 16, 2021 Introduction Background Chitons or polyplacophores constitute a of the eight classes A general synthesis about the current knowledge of the of the filum Mollusca. They are generally small, flattened and shallow water polyplacophoran molluscs (chitons) occurring elongated animals, provided with eight (8) overlapping dorsal in the Southern Caribbean of Venezuela is presented, involving plates or valves, bordered by a belt of scales and/or calcareous bibliographical revision/database (summarized informations [1] spicules formed by the mantle, called perinotum. The cavity of the from the “Northeast” region, coastal & insular, and [2] from “Los mantle encloses the foot, which is expands forming a large sole, Roques Archipelago Marine National Park”), and historical rescue which not only serves for the locomotion but also to maintain firm of informations (field notebooks) which includes inedit “personal -

122-2^0 Studies on the Common Rocky Egyptian Chiton
J, mar. biol Ass. India, 199^, 34 (1 & 2): 122-2^0 STUDIES ON THE COMMON ROCKY EGYPTIAN CHITON ACANTHOPLEURA GEMMATA (MOLLUSCA : POLYPLACOPHORA) IN THE NORTHWESTERN RED SEA FATHEY EL-SAYED SOUMAN AND Trro NAEM HABIB Department of Zoology, Faculty of Science, Sohag, Assiut University, Egypt ABSTRAOr In the present study on Acanthopleura gemmata (Blainville, 1825), rectification of the species name, its distribution and abundance in the northwestern part of the Red Sea, and the intraspeciflc variation within intermittent subpopulations have been achieved. Within each natural population, two forms, one banded and the other non^banded have been distinguished. The main differences in shell characters, girdle elements and radula features of the two forms have been investigated. INTRODUCTION In this study the distribution and abundance of the two forms along the Egyptian coastline THE POLYPLACOPHORAN Acanthopleura gemmcta has been recorded and morphometric data is one of the commonest in the rocky intertidal on specimens from several populations including area in the Indo-Paciflc province (Ferreira, the two forms, have been obtained to evaluate, 1986). Several studies on polyplacophorans if these forms are intraspeciflc varieties or have been carried out on the coastline of they are distinctly two dififerent species. the Red Sea (Fretter, 1937; Gunnar and Rupert, 1981 ; Iredale and Hull, 1923, The authors wish to express their gratitudes 1927). In these studies ten species of chitons to Prof. Dr. Ahmed A. El-Samahy, Vice- have been recorded, but their identification and President of Assiut University (Sohag), description were to some extent poor, as most Dr. Mohammed M. Ibrahim, Dean of Faculty of the authors agreed that the largest and of Science (Sohag) for providing facilities most common spscies are those related to genus during the course of this study. -

Proceedings of the United States National Museum
PROCEEDINGS OF UNITED STATES NATIONAL MUSEUM. 281 AVERAGE OF THE SPECIMENS. Length of head in total length without caudal (times) 4.62 luterorbital area in total length without caudal (times) 43 Snout in total length without caudal (times) 17 Upper jaw in total length without caudal (times) 14. 05 Mandible in total length Avithout caudal (times) - 11 Distance of dorsal from snout in total length without caudal (times) 4. 73 Base of dorsal in total length without caudal (times) 1. 26 Distance of anal from snout in total length without caudal (times) 2. 17 Base of anal in total length withoiit caudal (times) 1.84 Distance of pectoral snout total length without caudal (times) 4. 51 from in , Length of pectoral in total length without caudal (times) 5. 95 Distance of ventral from snout in total length without caudal (times) 4. 79 Length of ventral in total length without caudal (times) 13. 74 Branchiostegals VI Dorsal rays 48-50 Anal rays 33-37 Caudal rays 21-22 Pectoral rays 15 Ventral rays 3 U. S. Natioxal Museum, Washington, December 4, 1878. KEB'OUTT O.'V THE I^IMB»ETS AIV13 CMSTON.S OF THE ABLiASKAIV AIVO AKCTUt; KEGBOIVS, "WQTH I5E(^CBIIff»TII©^S ©IF GEIVEKA AlVD SPE- CIE.^ BELiUEVED TO BE IVEW. By ^W, IS. DAJLL. The followiug report has been drawn up chiefly from material collected in Alaska from 1865 to 1874 inclusive, but includes references to the few Arctic or northern species which are not common to Alaskan waters. The northwest coast of America, which I have already stated I have reason to think is the original center of distribution for the group of Doeo- glossa, at least of the littoral forms, is unquestionably the richest field where these animals may be found. -
Zootaxa,A Modern Look at the Animal Tree of Life
Zootaxa 1668: 61–79 (2007) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ ZOOTAXA Copyright © 2007 · Magnolia Press ISSN 1175-5334 (online edition) A modern look at the Animal Tree of Life* GONZALO GIRIBET1, CASEY W. DUNN2, GREGORY D. EDGECOMBE3, GREG W. ROUSE4 1Department of Organismic and Evolutionary Biology & Museum of Comparative Zoology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, USA, [email protected] 2Department of Ecology and Evolutionary Biology, Brown University, Providence, 80 Waterman Street, RI 02912, USA, [email protected] 3Department of Palaeontology, The Natural History Museum, Cromwell Road, London, SW7 5BD, UK, [email protected] 4Scripps Institution of Oceanography, University of California San Diego, 9500 Gilman Drive #0202, La Jolla, CA 92093, USA, [email protected] *In: Zhang, Z.-Q. & Shear, W.A. (Eds) (2007) Linnaeus Tercentenary: Progress in Invertebrate Taxonomy. Zootaxa, 1668, 1–766. Table of contents Abstract . .61 The setting . 62 The Animal Tree of Life—molecules and history . .62 The Animal Tree of Life—morphology and new developments . 63 Recent consensus on the Animal Tree of Life . 65 The base of the animal tree . .68 Bilateria . .72 Protostomia-Deuterostomia . .72 The Future of the Animal Tree of Life . .73 Acknowledgements . .73 References . 73 Abstract The phylogenetic interrelationships of animals (Metazoa) have been elucidated by refined systematic methods and by new techniques, notably from molecular biology. In parallel with the strong molecular focus of contemporary metazoan phylogenetics, morphology has advanced with the introduction of new approaches, such as confocal laser scanning microscopy and cell-labelling in the study of embryology.