A Metapopulation Model for Whale-Fall Specialists: the Largest Whales Are Essential to Prevent Species Extinctions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Role of Cetaceans in Ecosystem Functioning
WORKSHOP REPORT Role of Cetaceans in Ecosystem Functioning: Defining Marine Conservation Policies in the 21st Century 28th International Congress for Conservation Biology Society for Conservation Biology Workshop Report Role of Cetaceans in Ecosystem Functioning: Defining Marine Conservation Policies in the 21st Century 28th International Congress for Conservation Biology Society for Conservation Biology 26 July 2017, Cartagena, Colombia Room Barahona 1, Cartagena Convention Center www.ccc-chile.org www.icb.org.ar www.whales.org www.oceancare.org www.hsi.org csiwhalesalive.org www.nrdc.org www.minrel.gob.cl www.belgium.be For centuries, the great whales (baleen whales and the scientists, but to ecological economists (who ascribe finan- sperm whale) and other cetaceans1 (small whales, dolphins cial values to ecological functions) and to and porpoises) were valued almost exclusively for their oil policymakers concerned with conserving biodiversity. and meat. Widespread commercial hunting reduced great These services confirm what the public, since the early whale numbers by as much as 90 percent, with some ‘Save the Whale’ movement in the 1970s, has always un- populations being hunted to extinction. derstood; cetaceans are special. In recent decades, changing attitudes toward protecting The global implications of the significant contributions of wildlife and the natural world and the growth of ecotourism cetaceans “to ecosystem functioning that are beneficial for provided new cultural and non-extractive economic values the natural environment and people” were first formally for these marine mammals. acknowledged in 2016 when the International Whaling Commission (IWC) adopted a resolution on Cetaceans and Today, whale watching is worth more than $2 billion annu- Their Contributions to Ecosystem Functioning2. -

Deep-Sea Biology
Deep-Sea Biology (OCN430) - Syllabus Fall 2017 Instructors: Jeff Drazen, office MSB605, [email protected], 956-6567 Craig Smith, office MSB617, [email protected], 956-7776 T TH 12:00-1:15 POST708 Syllabus schedule subject to change Course Goals – The deep sea is the largest living space on the planet. Its inhabitants are varied and its communities are often complex, adapted to the particular characteristics of their habitat. This course will cover the major topics in the field, such as bentho-pelagic coupling, depth zonation, energetics, diversity, ecosystem function, adaptations, and the ecology of major habitats. The last portion of the course will deal with anthropogenic threats such as deep-sea fisheries, mining and global climate change. Its goal is to provide you with a basic understanding of what we know (and don’t know) about the biology, ecology and biodiversity of deep-sea ecosystems, the methods used in the field, and it will create a forum for discussion of the major current questions and recent exciting discoveries. Course Structure – After each lecture (or pair of lectures) students will lead a discussion session. The lectures will present the basics of the topics. The discussions will be based on assigned readings (primarily current scientific papers), allowing the class to explore the controversies, implications of recent findings, and highlight future directions for research. Student Learning Outcomes – At the end of this course you will be able to: 1) Describe the co-varying effects of temperature, pressure, oxygen and light levels on the adaptations of deep-sea organisms. 2) Evaluate the influence of variables co-varying with depth on communities, populations, and species. -

The First Record of the Genus Lamellibrachia (Siboglinidae
J. Earth Syst. Sci. (2021) 130:94 Ó Indian Academy of Sciences https://doi.org/10.1007/s12040-021-01587-1 (0123456789().,-volV)(0123456789().,-volV) The Brst record of the genus Lamellibrachia (Siboglinidae) tubeworm along with associated organisms in a chemosynthetic ecosystem from the Indian Ocean: A report from the Cauvery–Mannar Basin 1, 1 1 1 AMAZUMDAR *, P DEWANGAN ,APEKETI ,FIROZ BADESAAB , 1,5 1,6 1 1,6 MOHD SADIQUE ,KALYANI SIVAN ,JITTU MATHAI ,ANKITA GHOSH , 1,6 1,5 2 1,6 1 AZATALE ,SPKPILLUTLA ,CUMA ,CKMISHRA ,WALSH FERNANDES , 3 4 ASTHA TYAGI and TANOJIT PAUL 1Gas Hydrate Research Group, CSIR-National Institute of Oceanography, Dona Paula, Goa 403 004, India. 2Kerala University of Fisheries and Ocean Studies, Kochi, Kerala 682 506, India. 3K.J. Somaiya College of Science and Commerce, University of Mumbai, Mumbai, Maharashtra 400 077, India. 4Manipal Institute of Technology, Manipal, Karnataka 576 104, India. 5School of Earth, Ocean, and Atmospheric Sciences, Goa University, Taleigao Plateau, Goa 403 001, India. 6Academy of ScientiBc and Innovative Research (AcSIR), Ghaziabad, Uttar Pradesh 201 002, India. *Corresponding author. e-mail: [email protected] MS received 2 October 2020; revised 23 January 2021; accepted 25 January 2021 Here, we report for the Brst time, the genus Lamellibrachia tubeworm and associated chemosynthetic ecosystem from a cold-seep site in the Indian Ocean. The discovery of cold-seep was made oA the Cauvery–Mannar Basin onboard ORV Sindhu Sadhana (SSD-070; 13th to 22nd February 2020). The chemosymbiont bearing polychaete worm is also associated with squat lobsters (Munidposis sp.) and Gastropoda belonging to the family Buccinidae. -

Phylogenomics of Tubeworms (Siboglinidae, Annelida) and Comparative Performance of Different Reconstruction Methods
Zoologica Scripta Phylogenomics of tubeworms (Siboglinidae, Annelida) and comparative performance of different reconstruction methods YUANNING LI,KEVIN M. KOCOT,NATHAN V. WHELAN,SCOTT R. SANTOS,DAMIEN S. WAITS, DANIEL J. THORNHILL &KENNETH M. HALANYCH Submitted: 28 January 2016 Li, Y., Kocot, K.M., Whelan, N.V., Santos, S.R., Waits, D.S., Thornhill, D.J. & Halanych, Accepted: 18 June 2016 K.M. (2016). Phylogenomics of tubeworms (Siboglinidae, Annelida) and comparative perfor- doi:10.1111/zsc.12201 mance of different reconstruction methods. —Zoologica Scripta, 00: 000–000. Deep-sea tubeworms (Annelida, Siboglinidae) represent dominant species in deep-sea chemosynthetic communities (e.g. hydrothermal vents and cold methane seeps) and occur in muddy sediments and organic falls. Siboglinids lack a functional digestive tract as adults, and they rely on endosymbiotic bacteria for energy, making them of evolutionary and physi- ological interest. Despite their importance, inferred evolutionary history of this group has been inconsistent among studies based on different molecular markers. In particular, place- ment of bone-eating Osedax worms has been unclear in part because of their distinctive biol- ogy, including harbouring heterotrophic bacteria as endosymbionts, displaying extreme sexual dimorphism and exhibiting a distinct body plan. Here, we reconstructed siboglinid evolutionary history using 12 newly sequenced transcriptomes. We parsed data into three data sets that accommodated varying levels of missing data, and we evaluate effects of miss- ing data on phylogenomic inference. Additionally, several multispecies-coalescent approaches and Bayesian concordance analysis (BCA) were employed to allow for a compar- ison of results to a supermatrix approach. Every analysis conducted herein strongly sup- ported Osedax being most closely related to the Vestimentifera and Sclerolinum clade, rather than Frenulata, as previously reported. -
SDP PICOC Template
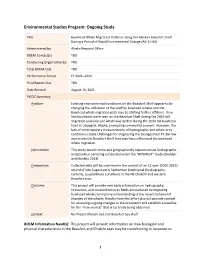
Environmental Studies Program: Ongoing Study Title Bowhead Whale Migration Patterns along the Alaskan Beaufort Shelf During a Period of Rapid Environmental Change (AK-21-04) Administered by Alaska Regional Office BOEM Contact(s) TBD Conducting Organization(s) TBD Total BOEM Cost TBD Performance Period FY 2021–2024 Final Report Due TBD Date Revised August 19, 2021 PICOC Summary Problem Evolving environmental conditions on the Beaufort Shelf appear to be changing the utilization of the shelf by bowhead whales and the bowhead whale migration path may be shifting farther offshore. Very few bowheads were seen on the Beaufort Shelf during the 2019 fall migration and only one whale was landed during the 2019 fall bowhead hunt at Utqiaġvik, Alaska, prompting community concern. However, the lack of contemporary measurements of hydrographic and whale prey conditions create challenges for diagnosing the changes near Pt. Barrow and across the Beaufort Shelf that may have influenced the bowhead whale migration. Intervention This study would renew and geographically expand annual hydrographic and plankton sampling conducted under the “BOWFEST” study (Shelden and Mocklin 2013). Comparison Collected data will be examined in the context of an 11-year (2005-2015) record of late August-early September biophysical (hydrography, currents, zooplankton) conditions in the NE Chukchi and western Beaufort seas. Outcome This project will provide new basic information on hydrography, circulation, and zooplankton prey fields encountered by migrating bowhead whales to improve understanding of the recent behavioral changes of the whales. Results from this effort also will provide context for assessing ongoing changes to the ecosystem and establish a baseline for the “new normal” that is currently being observed. -

Articles and Detrital Matter
Biogeosciences, 7, 2851–2899, 2010 www.biogeosciences.net/7/2851/2010/ Biogeosciences doi:10.5194/bg-7-2851-2010 © Author(s) 2010. CC Attribution 3.0 License. Deep, diverse and definitely different: unique attributes of the world’s largest ecosystem E. Ramirez-Llodra1, A. Brandt2, R. Danovaro3, B. De Mol4, E. Escobar5, C. R. German6, L. A. Levin7, P. Martinez Arbizu8, L. Menot9, P. Buhl-Mortensen10, B. E. Narayanaswamy11, C. R. Smith12, D. P. Tittensor13, P. A. Tyler14, A. Vanreusel15, and M. Vecchione16 1Institut de Ciencies` del Mar, CSIC. Passeig Mar´ıtim de la Barceloneta 37-49, 08003 Barcelona, Spain 2Biocentrum Grindel and Zoological Museum, Martin-Luther-King-Platz 3, 20146 Hamburg, Germany 3Department of Marine Sciences, Polytechnic University of Marche, Via Brecce Bianche, 60131 Ancona, Italy 4GRC Geociencies` Marines, Parc Cient´ıfic de Barcelona, Universitat de Barcelona, Adolf Florensa 8, 08028 Barcelona, Spain 5Universidad Nacional Autonoma´ de Mexico,´ Instituto de Ciencias del Mar y Limnolog´ıa, A.P. 70-305 Ciudad Universitaria, 04510 Mexico,` Mexico´ 6Woods Hole Oceanographic Institution, MS #24, Woods Hole, MA 02543, USA 7Integrative Oceanography Division, Scripps Institution of Oceanography, La Jolla, CA 92093-0218, USA 8Deutsches Zentrum fur¨ Marine Biodiversitatsforschung,¨ Sudstrand¨ 44, 26382 Wilhelmshaven, Germany 9Ifremer Brest, DEEP/LEP, BP 70, 29280 Plouzane, France 10Institute of Marine Research, P.O. Box 1870, Nordnes, 5817 Bergen, Norway 11Scottish Association for Marine Science, Scottish Marine Institute, Oban, -

Systematics, Evolution and Phylogeny of Annelida – a Morphological Perspective
Memoirs of Museum Victoria 71: 247–269 (2014) Published December 2014 ISSN 1447-2546 (Print) 1447-2554 (On-line) http://museumvictoria.com.au/about/books-and-journals/journals/memoirs-of-museum-victoria/ Systematics, evolution and phylogeny of Annelida – a morphological perspective GÜNTER PURSCHKE1,*, CHRISTOPH BLEIDORN2 AND TORSTEN STRUCK3 1 Zoology and Developmental Biology, Department of Biology and Chemistry, University of Osnabrück, Barbarastr. 11, 49069 Osnabrück, Germany ([email protected]) 2 Molecular Evolution and Animal Systematics, University of Leipzig, Talstr. 33, 04103 Leipzig, Germany (bleidorn@ rz.uni-leipzig.de) 3 Zoological Research Museum Alexander König, Adenauerallee 160, 53113 Bonn, Germany (torsten.struck.zfmk@uni- bonn.de) * To whom correspondence and reprint requests should be addressed. Email: [email protected] Abstract Purschke, G., Bleidorn, C. and Struck, T. 2014. Systematics, evolution and phylogeny of Annelida – a morphological perspective . Memoirs of Museum Victoria 71: 247–269. Annelida, traditionally divided into Polychaeta and Clitellata, is an evolutionary ancient and ecologically important group today usually considered to be monophyletic. However, there is a long debate regarding the in-group relationships as well as the direction of evolutionary changes within the group. This debate is correlated to the extraordinary evolutionary diversity of this group. Although annelids may generally be characterised as organisms with multiple repetitions of identically organised segments and usually bearing certain other characters such as a collagenous cuticle, chitinous chaetae or nuchal organs, none of these are present in every subgroup. This is even true for the annelid key character, segmentation. The first morphology-based cladistic analyses of polychaetes showed Polychaeta and Clitellata as sister groups. -

Whales, Wood and Kelp Islands in the Deep-Sea
ANGELO FRAGA BERNARDINO Whales, wood and kelp islands in the deep-sea: ecological succession and species overlap with other chemosynthetic habitats in the Californian continental slope (NE Pacific) Tese apresentada ao Instituto Oceanográfico da Universidade de São Paulo, como parte dos requisitos para obtenção do título de Doutor em Ciências, área de Oceanografia Biológica. Orientador: Prof. Dr. Paulo Yukio Gomes Sumida Co-orientador: Prof. Dr. Craig R. Smith São Paulo 2009 Universidade de São Paulo Instituto Oceanográfico Whales, wood and kelp islands in the deep-sea: ecological succession and species overlap with other chemosynthetic habitats in the Californian continental slope (NE Pacific) Angelo Fraga Bernardino Tese apresentada ao Instituto Oceanográfico da Universidade de São Paulo, como parte dos requisitos para obtenção do título de Doutor em Ciências, área de Oceanografia Biológica. Julgada em ___/___/_____ Prof.(a) Dr.(a) Conceito Prof.(a) Dr.(a) Conceito Prof.(a) Dr.(a) Conceito Prof.(a) Dr.(a) Conceito Prof.(a) Dr.(a) Conceito i INDEX List of Tables ……………………………………………………………….………… ii List of Figures ………………………………………………………………..………. iii Abstract ………………………………………………………………..……………… v Resumo ………………………………………………………………..……………… vi Chapter 1. Introduction ………………………………………………….…………….. 1 1.1. References ..………………………..….………………………………..…........ 6 Chapter 2. Macrofaunal succession in sediments around kelp and wood falls in the 9 deep NE Pacific and community overlap with other reducing habitats 2.1. Introduction ……………………………………………………………………. 10 2.2. Materials and methods ……………...........…………………………………….. 12 2.3. Results ………………………............................................................................. 19 2.4. Discussion …………………………….……………………………………….. 38 2.5. Conclusions …………….……………………………………………………… 46 2.6. References ……….…………………………………………………………….. 46 2.7. Supplementary material …………………………….…………………………. 55 Chapter 3. Infaunal community structure and succession during the sulfophilic stage 57 of a whale carcass in the deep NE Pacific 3.1. -

Ecology of Whale Falls at the Deep-Sea Floor
Oceanography and Marine Biology: an Annual Review 2003, 41, 311–354 © R.N. Gibson and R.J.A. Atkinson, Editors Taylor & Francis ECOLOGY OF WHALE FALLS AT THE DEEP-SEA FLOOR CRAIG R. SMITH1 & AMY R. BACO1,2 1Department of Oceanography, University of Hawaii at Manoa, 1000 Pope Road, Honolulu, HI, 96822, USA e-mail: [email protected] 2present address: Biology Department, Woods Hole Oceanographic Institution, Woods Hole, MA 02543, USA e-mail: [email protected] Abstract The falls of large whales (30–160t adult body weight) yield massive pulses of labile organic matter to the deep-sea floor. While scientists have long speculated on the ecological roles of such concentrated food inputs, observations have accumulated since the 1850s to suggest that deep-sea whale falls support a widespread, characteristic fauna. Interest in whale- fall ecology heightened with the discovery in 1989 of a chemoautotrophic assemblage on a whale skeleton in the northeast Pacific; related communities were soon reported from whale falls in other bathyal and abyssal Pacific and Atlantic sites, and from 30mya (million years ago) in the northeast Pacific fossil record. Recent time-series studies of natural and implanted deep- sea whale falls off California, USA indicate that bathyal carcasses pass through at least three successional stages: (1) a mobile-scavenger stage lasting months to years, during which aggregations of sleeper sharks, hagfish, rat-tails and invertebrate scavengers remove whale soft tissue at high rates (40–60kgdϪ1); (2) an enrichment opportunist stage (duration of months to years) during which organi- cally enriched sediments and exposed bones are colonised by dense assemblages (up to 40000mϪ2) of opportunistic polychaetes and crustaceans; (3) a sulphophilic (“or sulphur-loving”) stage lasting for decades, during which a large, species-rich, trophically complex assemblage lives on the skeleton as it emits sul- phide from anaerobic breakdown of bone lipids; this stage includes a chemoau- totrophic component deriving nutrition from sulphur-oxidising bacteria. -

Siboglinidae, Annelida) from Cold Seeps Off New Zealand, and an Assessment of Its Presence in the Western Pacific Ocean Marina F
McCowin et al. Marine Biodiversity Records (2019) 12:10 https://doi.org/10.1186/s41200-019-0169-2 MARINERECORD Open Access A new record of Lamellibrachia columna (Siboglinidae, Annelida) from cold seeps off New Zealand, and an assessment of its presence in the western Pacific Ocean Marina F. McCowin1*, Ashley A. Rowden2 and Greg W. Rouse1 Abstract Lamellibrachia columna Southward was originally described from hydrothermal vents of the Lau Basin, between Fiji and Tonga. This study utilizes phylogenetic and morphological analyses to confirm the collection of Lamellibrachia columna from cold seeps on the Hikurangi Margin off New Zealand, thereby extending its geographic range southward by approximately 1900 km. We also propose, based on molecular evidence, that specimens previously reported from vents in the Nankai Trough, Japan and seeps off southern and eastern Japan are L. columna. Furthermore, we suggest that Lamellibrachia sagami Kobayashi et al. described from cold seeps off southern and eastern Japan is a junior synonym of Lamellibrachia columna. Our work confirms that L. columna is found at two types of chemosynthetic habitat over a wide geographic range in the western Pacific Ocean. Keywords: Vestimentifera, Deep sea, Pacific Ocean, Phylogeny, Synonymy Background to be described from the southwestern Pacific, at 1870 m Members of Vestimentifera (originally introduced by in an area of diffuse venting at the Lau Back-arc Basin Jones (1985), and now one of four informal lineages (Southward 1991). Since its initial discovery, L. columna within Siboglinidae Caullery, 1914 [Hilário et al. 2011; has been reported at diffuse vents from the southwestern Pleijel et al. 2009]) live in chemosynthetic environments Pacific back-arc basin at Lau from 1832 to 1914 m such as hydrothermal vents (Bright and Lallier 2010; (Black et al. -

Black Spicules from a New Interstitial Opheliid Polychaete Thoracophelia Minuta Sp
www.nature.com/scientificreports OPEN Black spicules from a new interstitial opheliid polychaete Thoracophelia minuta sp. nov. (Annelida: Opheliidae) Naoto Jimi1*, Shinta Fujimoto2, Mami Takehara1 & Satoshi Imura1,3 The phylum Annelida exhibits high morphological diversity coupled with its extensive ecological diversity, and the process of its evolution has been an attractive research subject for many researchers. Its representatives are also extensively studied in felds of ecology and developmental biology and important in many other biology related disciplines. The study of biomineralisation is one of them. Some annelid groups are well known to form calcifed tubes but other forms of biomineralisation are also known. Herein, we report a new interstitial annelid species with black spicules, Thoracophelia minuta sp. nov., from Yoichi, Hokkaido, Japan. Spicules are minute calcium carbonate inclusions found across the body and in this new species, numerous black rod-like inclusions of calcium-rich composition are distributed in the coelomic cavity. The new species can be distinguished from other known species of the genus by these conspicuous spicules, shape of branchiae and body formula. Further, the new species’ body size is apparently smaller than its congeners. Based on our molecular phylogenetic analysis using 18S and 28S sequences, we discuss the evolutionary signifcance of the new species’ spicules and also the species’ progenetic origin. Annelida is one of the most ecologically and morphologically diverse group of animals known from both marine and terrestrial environments. Several groups are highly specialised with distinct ecological niches such as intersti- tial, parasitic, pelagic, or chemosynthetic zones 1. Like many other animal phyla 2–6, annelids are known to produce biominerals2. -

Deep-Sea Whale Fall Fauna from the Atlantic Resembles That of the Pacific Ocean Received: 03 August 2015 Paulo Y
www.nature.com/scientificreports OPEN Deep-sea whale fall fauna from the Atlantic resembles that of the Pacific Ocean Received: 03 August 2015 Paulo Y. G. Sumida1, Joan M. Alfaro-Lucas1, Mauricio Shimabukuro1, Hiroshi Kitazato2, Accepted: 08 February 2016 Jose A. A. Perez3, Abilio Soares-Gomes4, Takashi Toyofuku2, Andre O. S. Lima3, Koichi Ara5 & Published: 24 February 2016 Yoshihiro Fujiwara2 Whale carcasses create remarkable habitats in the deep-sea by producing concentrated sources of organic matter for a food-deprived biota as well as places of evolutionary novelty and biodiversity. Although many of the faunal patterns on whale falls have already been described, the biogeography of these communities is still poorly known especially from basins other than the NE Pacific Ocean. The present work describes the community composition of the deepest natural whale carcass described to date found at 4204 m depth on Southwest Atlantic Ocean with manned submersible Shinkai 6500. This is the first record of a natural whale fall in the deep Atlantic Ocean. The skeleton belonged to an Antarctic Minke whale composed of only nine caudal vertebrae, whose degradation state suggests it was on the bottom for 5–10 years. The fauna consisted mainly of galatheid crabs, a new species of the snail Rubyspira and polychaete worms, including a new Osedax species. Most of the 41 species found in the carcass are new to science, with several genera shared with NE Pacific whale falls and vent and seep ecosystems. This similarity suggests the whale-fall fauna is widespread and has dispersed in a stepping stone fashion, deeply influencing its evolutionary history.