Key Concepts -- Lecture 5 IB 168: Spring 2009 Polypodiales
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Genetic Differentiation and Polyploid Formation Within the Cryptogramma Crispa Complex (Polypodiales: Pteridaceae)
Turkish Journal of Botany Turk J Bot (2016) 40: 231-240 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1501-54 Genetic differentiation and polyploid formation within the Cryptogramma crispa complex (Polypodiales: Pteridaceae) Jordan METZGAR*, Mackenzie STAMEY, Stefanie ICKERT-BOND Herbarium (ALA), University of Alaska Museum of the North and Department of Biology and Wildlife, University of Alaska Fairbanks, Fairbanks, AK, USA Received: 28.01.2015 Accepted/Published Online: 14.07.2015 Final Version: 08.04.2016 Abstract: The tetraploid fern Cryptogramma crispa (L.) R.Br. ex Hook. is distributed across alpine and high latitude regions of Europe and western Asia and is sympatric with the recently described octoploid C. bithynica S.Jess., L.Lehm. & Bujnoch in north-central Turkey. Our analysis of a 6-region plastid DNA sequence dataset comprising 39 accessions of Cryptogramma R.Br., including 14 accessions of C. crispa and one accession of C. bithynica, revealed a deep genetic division between the accessions of C. crispa from western, northern, and central Europe and the accessions of C. crispa and C. bithynica from Turkey and the Caucasus Mountains. This legacy likely results from Pleistocene climate fluctuations and appears to represent incipient speciation between the eastern and western clades. These plastid DNA sequence data also demonstrate that the western clade of C. crispa, specifically the western Asian clade, is the maternal progenitor of C. bithynica. Our analysis of DNA sequence data from the biparentally inherited nuclear locus gapCp supports an autopolyploid origin of C. bithynica, with C. crispa as the sole progenitor. Key words: Cryptogramma, ferns, autopolyploidy, phylogeography, glacial refugium 1. -

Monilophyte Mitochondrial Rps1 Genes Carry a Unique Group II Intron That Likely Originated from an Ancient Paralog in Rpl2
Downloaded from rnajournal.cshlp.org on September 26, 2021 - Published by Cold Spring Harbor Laboratory Press Monilophyte mitochondrial rps1 genes carry a unique group II intron that likely originated from an ancient paralog in rpl2 NILS KNIE, FELIX GREWE,1 and VOLKER KNOOP Abteilung Molekulare Evolution, IZMB–Institut für Zelluläre und Molekulare Botanik, Universität Bonn, D-53115 Bonn, Germany ABSTRACT Intron patterns in plant mitochondrial genomes differ significantly between the major land plant clades. We here report on a new, clade-specific group II intron in the rps1 gene of monilophytes (ferns). This intron, rps1i25g2, is strikingly similar to rpl2i846g2 previously identified in the mitochondrial rpl2 gene of seed plants, ferns, and the lycophyte Phlegmariurus squarrosus. Although mitochondrial ribosomal protein genes are frequently subject to endosymbiotic gene transfer among plants, we could retrieve the mitochondrial rps1 gene in a taxonomically wide sampling of 44 monilophyte taxa including basal lineages such as the Ophioglossales, Psilotales, and Marattiales with the only exception being the Equisetales (horsetails). Introns rps1i25g2 and rpl2i846g2 were likewise consistently present with only two exceptions: Intron rps1i25g2 is lost in the genus Ophioglossum and intron rpl2i846g2 is lost in Equisetum bogotense. Both intron sequences are moderately affected by RNA editing. The unprecedented primary and secondary structure similarity of rps1i25g2 and rpl2i846g2 suggests an ancient retrotransposition event copying rpl2i846g2 into rps1, for which we suggest a model. Our phylogenetic analysis adding the new rps1 locus to a previous data set is fully congruent with recent insights on monilophyte phylogeny and further supports a sister relationship of Gleicheniales and Hymenophyllales. Keywords: group II intron; RNA editing; intron transfer; reverse splicing; intron loss; monilophyte phylogeny INTRODUCTION and accordingly the genome sizes of most free-living bacteria, are an example for the latter (Ward et al. -

Horizontal Transfer of an Adaptive Chimeric Photoreceptor from Bryophytes to Ferns
Horizontal transfer of an adaptive chimeric photoreceptor from bryophytes to ferns Fay-Wei Lia,1, Juan Carlos Villarrealb, Steven Kellyc, Carl J. Rothfelsd, Michael Melkoniane, Eftychios Frangedakisc, Markus Ruhsamf, Erin M. Sigela, Joshua P. Derg,h, Jarmila Pittermanni, Dylan O. Burgej, Lisa Pokornyk, Anders Larssonl, Tao Chenm, Stina Weststrandl, Philip Thomasf, Eric Carpentern, Yong Zhango, Zhijian Tiano, Li Cheno, Zhixiang Yano, Ying Zhuo, Xiao Suno, Jun Wango, Dennis W. Stevensonp, Barbara J. Crandall-Stotlerq, A. Jonathan Shawa, Michael K. Deyholosn, Douglas E. Soltisr,s,t, Sean W. Grahamu, Michael D. Windhama, Jane A. Langdalec, Gane Ka-Shu Wongn,o,v,1, Sarah Mathewsw, and Kathleen M. Pryera aDepartment of Biology, Duke University, Durham, NC 27708; bSystematic Botany and Mycology, Department of Biology, University of Munich, 80638 Munich, Germany; cDepartment of Plant Sciences, University of Oxford, Oxford OX1 3RB, United Kingdom; dDepartment of Zoology, University of British Columbia, Vancouver, BC, Canada V6T 1Z4; eBotany Department, Cologne Biocenter, University of Cologne, 50674 Cologne, Germany; fRoyal Botanic Garden Edinburgh, Edinburgh EH3 5LR, Scotland; gDepartment of Biology and hHuck Institutes of the Life Sciences, Pennsylvania State University, University Park, PA 16802; iDepartment of Ecology and Evolutionary Biology, University of California, Santa Cruz, CA 95064; jCalifornia Academy of Sciences, San Francisco, CA 94118; kReal Jardín Botánico, 28014 Madrid, Spain; lSystematic Biology, Evolutionary Biology Centre, -

Guidance Document Pohakuloa Training Area Plant Guide
GUIDANCE DOCUMENT Recovery of Native Plant Communities and Ecological Processes Following Removal of Non-native, Invasive Ungulates from Pacific Island Forests Pohakuloa Training Area Plant Guide SERDP Project RC-2433 JULY 2018 Creighton Litton Rebecca Cole University of Hawaii at Manoa Distribution Statement A Page Intentionally Left Blank This report was prepared under contract to the Department of Defense Strategic Environmental Research and Development Program (SERDP). The publication of this report does not indicate endorsement by the Department of Defense, nor should the contents be construed as reflecting the official policy or position of the Department of Defense. Reference herein to any specific commercial product, process, or service by trade name, trademark, manufacturer, or otherwise, does not necessarily constitute or imply its endorsement, recommendation, or favoring by the Department of Defense. Page Intentionally Left Blank 47 Page Intentionally Left Blank 1. Ferns & Fern Allies Order: Polypodiales Family: Aspleniaceae (Spleenworts) Asplenium peruvianum var. insulare – fragile fern (Endangered) Delicate ENDEMIC plants usually growing in cracks or caves; largest pinnae usually <6mm long, tips blunt, uniform in shape, shallowly lobed, 2-5 lobes on acroscopic side. Fewer than 5 sori per pinna. Fronds with distal stipes, proximal rachises ocassionally proliferous . d b a Asplenium trichomanes subsp. densum – ‘oāli’i; maidenhair spleenwort Plants small, commonly growing in full sunlight. Rhizomes short, erect, retaining many dark brown, shiny old stipe bases.. Stipes wiry, dark brown – black, up to 10cm, shiny, glabrous, adaxial surface flat, with 2 greenish ridges on either side. Pinnae 15-45 pairs, almost sessile, alternate, ovate to round, basal pinnae smaller and more widely spaced. -

Millerocaulis Tekelili Sp. Nov., a New Species of Osmundalean Fern from the Aptian Cerro Negro Formation (Antarctica) Ezequiel I
This article was downloaded by: [INASP - Pakistan ] On: 28 July 2011, At: 10:58 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Alcheringa: An Australasian Journal of Palaeontology Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/talc20 Millerocaulis tekelili sp. nov., a new species of osmundalean fern from the Aptian Cerro Negro Formation (Antarctica) Ezequiel I. Vera Available online: 27 Jul 2011 To cite this article: Ezequiel I. Vera (2011): Millerocaulis tekelili sp. nov., a new species of osmundalean fern from the Aptian Cerro Negro Formation (Antarctica), Alcheringa: An Australasian Journal of Palaeontology, DOI:10.1080/03115518.2011.576541 To link to this article: http://dx.doi.org/10.1080/03115518.2011.576541 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.tandfonline.com/page/terms-and-conditions This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan, sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. -

(Polypodiales) Plastomes Reveals Two Hypervariable Regions Maria D
Logacheva et al. BMC Plant Biology 2017, 17(Suppl 2):255 DOI 10.1186/s12870-017-1195-z RESEARCH Open Access Comparative analysis of inverted repeats of polypod fern (Polypodiales) plastomes reveals two hypervariable regions Maria D. Logacheva1, Anastasiya A. Krinitsina1, Maxim S. Belenikin1,2, Kamil Khafizov2,3, Evgenii A. Konorov1,4, Sergey V. Kuptsov1 and Anna S. Speranskaya1,3* From Belyaev Conference Novosibirsk, Russia. 07-10 August 2017 Abstract Background: Ferns are large and underexplored group of vascular plants (~ 11 thousands species). The genomic data available by now include low coverage nuclear genomes sequences and partial sequences of mitochondrial genomes for six species and several plastid genomes. Results: We characterized plastid genomes of three species of Dryopteris, which is one of the largest fern genera, using sequencing of chloroplast DNA enriched samples and performed comparative analysis with available plastomes of Polypodiales, the most species-rich group of ferns. We also sequenced the plastome of Adianthum hispidulum (Pteridaceae). Unexpectedly, we found high variability in the IR region, including duplication of rrn16 in D. blanfordii, complete loss of trnI-GAU in D. filix-mas, its pseudogenization due to the loss of an exon in D. blanfordii. Analysis of previously reported plastomes of Polypodiales demonstrated that Woodwardia unigemmata and Lepisorus clathratus have unusual insertions in the IR region. The sequence of these inserted regions has high similarity to several LSC fragments of ferns outside of Polypodiales and to spacer between tRNA-CGA and tRNA-TTT genes of mitochondrial genome of Asplenium nidus. We suggest that this reflects the ancient DNA transfer from mitochondrial to plastid genome occurred in a common ancestor of ferns. -

Osmundaceae Royal Fern Family
Osmundaceae Royal Fern Family A family of large coarse plants, these are our largest pteridophyte species. Stipes arise from rhizomes Page | 70 covered in persistent frond bases. Stipes are pubescent but without scales, bearing stipules at their bases. Fertile fronds may be separate from vegetative fronds, or fronds divided between sterile and fertile portions. Sporangia are not arranged in sori, but in short stalked clusters. Osmunda L. royal ferns A single genus finds its way to eastern North America; three species grow in Nova Scotia. All have pinnate or bipinnate fronds. Key to species A. Fronds twice-pinnate; fertile pinnae carried at the top. Osmunda regalis aa. Fronds pinnate; sterile fronds deeply divided, lobes nearly entire. B B. Fertile pinnae near the middle of sterile fronds; no woolly O. claytoniana pubescence at the base of pinnae. bb. Fertile fronds on separate stipes; pinnae with a tuft of pubescence O. cinnamomea in the axils. Osmunda cinnamomea L. Cinnamon Fern; osmonde cannelle A common species, its fertile fronds are cinnamon– coloured, comprising minute sporangia appressed to the axis. Sterile fronds have pinnae subtended by clusters of pale pubescence. Fiddleheads emerge covered in pale or white coloured tomentum. Spores in spring. Grows in poorly-drained soils as found in bogs, swamps, coastal barrens and even low-lying pastures and wet conifer woods. 1-9 Osmundaceae Photo by Sean Blaney Very common throughout the province. Elsewhere from NF to ON, south to TX and FL; South America; Eurasia. Page | 71 Photo by David Mazerolle Osmunda claytoniana L. Interrupted Fern; osmonde de Clayton Fronds resemble the species above, but with both fertile and sterile pinnae on the same stipe in this species. -

Taxonomic, Phylogenetic, and Functional Diversity of Ferns at Three Differently Disturbed Sites in Longnan County, China
diversity Article Taxonomic, Phylogenetic, and Functional Diversity of Ferns at Three Differently Disturbed Sites in Longnan County, China Xiaohua Dai 1,2,* , Chunfa Chen 1, Zhongyang Li 1 and Xuexiong Wang 1 1 Leafminer Group, School of Life Sciences, Gannan Normal University, Ganzhou 341000, China; [email protected] (C.C.); [email protected] (Z.L.); [email protected] (X.W.) 2 National Navel-Orange Engineering Research Center, Ganzhou 341000, China * Correspondence: [email protected] or [email protected]; Tel.: +86-137-6398-8183 Received: 16 March 2020; Accepted: 30 March 2020; Published: 1 April 2020 Abstract: Human disturbances are greatly threatening to the biodiversity of vascular plants. Compared to seed plants, the diversity patterns of ferns have been poorly studied along disturbance gradients, including aspects of their taxonomic, phylogenetic, and functional diversity. Longnan County, a biodiversity hotspot in the subtropical zone in South China, was selected to obtain a more thorough picture of the fern–disturbance relationship, in particular, the taxonomic, phylogenetic, and functional diversity of ferns at different levels of disturbance. In 90 sample plots of 5 5 m2 along roadsides × at three sites, we recorded a total of 20 families, 50 genera, and 99 species of ferns, as well as 9759 individual ferns. The sample coverage curve indicated that the sampling effort was sufficient for biodiversity analysis. In general, the taxonomic, phylogenetic, and functional diversity measured by Hill numbers of order q = 0–3 indicated that the fern diversity in Longnan County was largely influenced by the level of human disturbance, which supports the ‘increasing disturbance hypothesis’. -

The Ferns and Their Relatives (Lycophytes)
N M D R maidenhair fern Adiantum pedatum sensitive fern Onoclea sensibilis N D N N D D Christmas fern Polystichum acrostichoides bracken fern Pteridium aquilinum N D P P rattlesnake fern (top) Botrychium virginianum ebony spleenwort Asplenium platyneuron walking fern Asplenium rhizophyllum bronze grapefern (bottom) B. dissectum v. obliquum N N D D N N N R D D broad beech fern Phegopteris hexagonoptera royal fern Osmunda regalis N D N D common woodsia Woodsia obtusa scouring rush Equisetum hyemale adder’s tongue fern Ophioglossum vulgatum P P P P N D M R spinulose wood fern (left & inset) Dryopteris carthusiana marginal shield fern (right & inset) Dryopteris marginalis narrow-leaved glade fern Diplazium pycnocarpon M R N N D D purple cliff brake Pellaea atropurpurea shining fir moss Huperzia lucidula cinnamon fern Osmunda cinnamomea M R N M D R Appalachian filmy fern Trichomanes boschianum rock polypody Polypodium virginianum T N J D eastern marsh fern Thelypteris palustris silvery glade fern Deparia acrostichoides southern running pine Diphasiastrum digitatum T N J D T T black-footed quillwort Isoëtes melanopoda J Mexican mosquito fern Azolla mexicana J M R N N P P D D northern lady fern Athyrium felix-femina slender lip fern Cheilanthes feei net-veined chain fern Woodwardia areolata meadow spike moss Selaginella apoda water clover Marsilea quadrifolia Polypodiaceae Polypodium virginanum Dryopteris carthusiana he ferns and their relatives (lycophytes) living today give us a is tree shows a current concept of the Dryopteridaceae Dryopteris marginalis is poster made possible by: { Polystichum acrostichoides T evolutionary relationships among Onocleaceae Onoclea sensibilis glimpse of what the earth’s vegetation looked like hundreds of Blechnaceae Woodwardia areolata Illinois fern ( green ) and lycophyte Thelypteridaceae Phegopteris hexagonoptera millions of years ago when they were the dominant plants. -
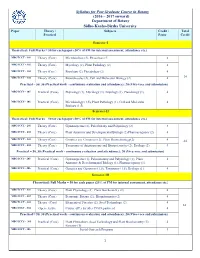
Syllabus for Post Graduate Course in Botany (2016 – 2017 Onward)
Syllabus for Post Graduate Course in Botany (2016 – 2017 onward) Department of Botany Sidho-Kanho-Birsha University Paper Theory / Subjects Credit / Total Practical Paper Credit Semester-I Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 101 Theory (Core) Microbiology (2), Phycology (2) 4 MBOTCCT - 102 Theory (Core) Mycology (2), Plant Pathology (2) 4 MBOTCCT - 103 Theory (Core) Bryology (2), Pteridology (2) 4 MBOTCCT - 104 Theory (Core) Biomolecules (2), Cell and Molecular Biology (2) 4 24 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 105 Practical (Core) Phycology (1), Mycology (1), Bryology (1), Pteridology (1). 4 MBOTCCS - 106 Practical (Core) Microbiology (1.5), Plant Pathology (1), Cell and Molecular 4 Biology (1.5). Semester-II Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 201 Theory (Core) Gymnosperms (2), Paleobotany and Palynology (2) 4 MBOTCCT - 202 Theory (Core) Plant Anatomy and Developmental Biology (2) Pharmacognosy (2) 4 MBOTCCT - 203 Theory (Core) Genetics and Genomics (2), Plant Biotechnology(2) 4 24 MBOTCCT - 204 Theory (Core) Taxonomy of Angiosperms and Biosystematics (2), Ecology (2) 4 Practical = 50, 30 (Practical work - continuous evaluation and attendance); 20 (Viva-voce and submission) MBOTCCS - 205 Practical (Core) Gymnosperms (1), Palaeobotany and Palynology (1), Plant 4 Anatomy & Developmental Biology (1), Pharmacognosy (1). MBOTCCS - 206 Practical (Core) Genetics and Genomics (1.5), Taxonomy (1.5), Ecology (1). 4 Semester-III Theoretical: Full Marks = 50 for each paper (20% of FM for internal assessment, attendance etc.) MBOTCCT - 301 Theory (Core) Plant Physiology (2), Plant Biochemistry (2) 4 MBOTCCT - 302 Theory (Core) Economic Botany (2), Bioinformatics (2) 4 MBOTCCT - 303 Theory (Core) Elements of Forestry (2), Seed Technology (2). -

Reprint Requests, Current Address: Dept
American Journal of Botany 88(6): 1118±1130. 2001. RBCL DATA REVEAL TWO MONOPHYLETIC GROUPS OF FILMY FERNS (FILICOPSIDA:HYMENOPHYLLACEAE)1 KATHLEEN M. PRYER,2,5 ALAN R. SMITH,3 JEFFREY S. HUNT,2 AND JEAN-YVES DUBUISSON4 2Department of Botany, The Field Museum of Natural History, 1400 S. Lake Shore Drive, Chicago, Illinois 60605-2496 USA; 3University Herbarium, 1001 Valley Life Sciences Building #2465, University of California, Berkeley, California 94720-2465 USA; and 4Laboratoire de PaleÂobotanique et PaleÂoeÂcologie, FR3-CNRS ``Institut d'E cologie,'' Universite Pierre et Marie Curie, 12 rue Cuvier, F-75005 Paris, France The ``®lmy fern'' family, Hymenophyllaceae, is traditionally partitioned into two principal genera, Trichomanes s.l. (sensu lato) and Hymenophyllum s.l., based upon sorus shape characters. This basic split in the family has been widely debated this past century and hence was evaluated here by using rbcL nucleotide sequence data in a phylogenetic study of 26 ®lmy ferns and nine outgroup taxa. Our results con®rm the monophyly of the family and provide robust support for two monophyletic groups that correspond to the two classical genera. In addition, we show that some taxa of uncertain af®nity, such as the monotypic genera Cardiomanes and Serpyllopsis, and at least one species of Microtrichomanes, are convincingly included within Hymenophyllum s.l. The tubular- or conical-based sorus that typi®es Trichomanes s.l. and Cardiomanes, the most basal member of Hymenophyllum s.l., is a plesiomorphic character state for the family. Tubular-based sori occurring in other members of Hymenophyllum s.l. are most likely derived independently and more than one time. -

Iop Newsletter 120
IOP NEWSLETTER 120 October 2019 CONTENTS Letter from the president Elections of IOP Executive Committee 2020 – Call for nominations Special issue in occasion of the 65th birthday of Hans Kerp Collection Spotlight: Cleveland Museum of Natural History Reflections from the ‘Earth Day’ 2019 Upcoming meetings IOP Logo: The evolution of plant architecture (© by A. R. Hemsley) 1 Letter from the president Greetings Members, This past quarter we have welcomed the publication Festschrifts celebrating our colleagues Hans Kerp (PalZ Paläontologische Zeitschrift, see below) and Gar Rothwell (International Journal of Plant Sciences v 180 nos. 7, 8). Thank you to the teams of editors and authors who have contributed to these issues illustrating the continuing strength of our discipline— and congratulations to Hans and Gar for inspiring such productivity by your admiring colleagues! Already three years have passed since our gathering for IOPC in Salvador, Brazil in 2016, and now planning is in full swing for next year’s meeting in Prague, 12-19 September 2020. This will be the 11th quadrennial meeting of IOP, held concurrently with the 15th International Palynological Conference. Please note the formal announcement in this newsletter (page 12, herein). Nominations for colleagues deserving of honorary membership are welcome at any time. As the submission of abstracts for IOPC/IPC presentations will be possible soon, we kindly remind the IOP Student Travel Awards. We will financially support about 5 to 7 PhD or MSc students in order to enable them to participate in the conference and present their research results in a talk. Recent PhD graduates will also qualify for these awards, if their completion was less than nine months prior to the time of the conference.