A Taxonomic Review of Ichthyosaurus from Somerset a Thesis Submitted To
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

A Mysterious Giant Ichthyosaur from the Lowermost Jurassic of Wales
A mysterious giant ichthyosaur from the lowermost Jurassic of Wales JEREMY E. MARTIN, PEGGY VINCENT, GUILLAUME SUAN, TOM SHARPE, PETER HODGES, MATT WILLIAMS, CINDY HOWELLS, and VALENTIN FISCHER Ichthyosaurs rapidly diversified and colonised a wide range vians may challenge our understanding of their evolutionary of ecological niches during the Early and Middle Triassic history. period, but experienced a major decline in diversity near the Here we describe a radius of exceptional size, collected at end of the Triassic. Timing and causes of this demise and the Penarth on the coast of south Wales near Cardiff, UK. This subsequent rapid radiation of the diverse, but less disparate, specimen is comparable in morphology and size to the radius parvipelvian ichthyosaurs are still unknown, notably be- of shastasaurids, and it is likely that it comes from a strati- cause of inadequate sampling in strata of latest Triassic age. graphic horizon considerably younger than the last definite Here, we describe an exceptionally large radius from Lower occurrence of this family, the middle Norian (Motani 2005), Jurassic deposits at Penarth near Cardiff, south Wales (UK) although remains attributable to shastasaurid-like forms from the morphology of which places it within the giant Triassic the Rhaetian of France were mentioned by Bardet et al. (1999) shastasaurids. A tentative total body size estimate, based on and very recently by Fischer et al. (2014). a regression analysis of various complete ichthyosaur skele- Institutional abbreviations.—BRLSI, Bath Royal Literary tons, yields a value of 12–15 m. The specimen is substantially and Scientific Institution, Bath, UK; NHM, Natural History younger than any previously reported last known occur- Museum, London, UK; NMW, National Museum of Wales, rences of shastasaurids and implies a Lazarus range in the Cardiff, UK; SMNS, Staatliches Museum für Naturkunde, lowermost Jurassic for this ichthyosaur morphotype. -

Mary Anning of Lyme Regis: 19Th Century Pioneer in British Palaeontology
Headwaters Volume 26 Article 14 2009 Mary Anning of Lyme Regis: 19th Century Pioneer in British Palaeontology Larry E. Davis College of St. Benedict / St. John's University, [email protected] Follow this and additional works at: https://digitalcommons.csbsju.edu/headwaters Part of the Geology Commons, and the Paleontology Commons Recommended Citation Davis, Larry E. (2009) "Mary Anning of Lyme Regis: 19th Century Pioneer in British Palaeontology," Headwaters: Vol. 26, 96-126. Available at: https://digitalcommons.csbsju.edu/headwaters/vol26/iss1/14 This Article is brought to you for free and open access by DigitalCommons@CSB/SJU. It has been accepted for inclusion in Headwaters by an authorized editor of DigitalCommons@CSB/SJU. For more information, please contact [email protected]. LARRY E. DAVIS Mary Anning of Lyme Regis 19th Century Pioneer in British Palaeontology Ludwig Leichhardt, a 19th century German explorer noted in a letter, “… we had the pleasure of making the acquaintance of the Princess of Palaeontology, Miss Anning. She is a strong, energetic spinster of about 28 years of age, tanned and masculine in expression …” (Aurousseau, 1968). Gideon Mantell, a 19th century British palaeontologist, made a less flattering remark when he wrote in his journal, “… sallied out in quest of Mary An- ning, the geological lioness … we found her in a little dirt shop with hundreds of specimens piled around her in the greatest disorder. She, the presiding Deity, a prim, pedantic vinegar looking female; shred, and rather satirical in her conversation” (Curwin, 1940). Who was Mary Anning, this Princess of Palaeontology and Geological Lioness (Fig. -

'Cenoceras Islands' in the Blue Lias Formation (Lower Jurassic)
FOSSIL IMPRINT • vol. 75 • 2019 • no. 1 • pp. 108–119 (formerly ACTA MUSEI NATIONALIS PRAGAE, Series B – Historia Naturalis) ‘CENOCERAS ISLANDS’ IN THE BLUE LIAS FORMATION (LOWER JURASSIC) OF WEST SOMERSET, UK: NAUTILID DOMINANCE AND INFLUENCE ON BENTHIC FAUNAS DAVID H. EVANS1, *, ANDY H. KING2 1 Stratigrapher, Natural England, Rivers House, East Quay, Bridgwater, Somerset, TA6 4YS UK; e-mail: [email protected]. 2 Director & Principal Geologist, Geckoella Ltd, Suite 323, 7 Bridge Street, Taunton, Somerset, TA1 1TG UK; e-mail: [email protected]. * corresponding author Evans, D. H., King, A. H. (2019): ‘Cenoceras islands’ in the Blue Lias Formation (Lower Jurassic) of West Somerset, UK: nautilid dominance and influence on benthic faunas. – Fossil Imprint, 75(1): 108–119, Praha. ISSN 2533-4050 (print), ISSN 2533-4069 (on-line). Abstract: Substantial numbers of the nautilid Cenoceras occur in a stratigraphically limited horizon within the upper part of the Lower Jurassic (Sinemurian Stage) Blue Lias Formation at Watchet on the West Somerset Coast (United Kingdom). Individual nautilid conchs are associated with clusters of encrusting organisms (sclerobionts) forming ‘islands’ that may have been raised slightly above the surrounding substrate. Despite the relatively large numbers of nautilid conchs involved, detailed investigation of their preservation suggests that their accumulation reflects a reduction in sedimentation rates rather than an influx of empty conches or moribund animals. Throughout those horizons in which nautilids are present in relative abundance, the remains of ammonites are subordinate or rare. The reason for this unclear, and preferential dissolution of ammonite conchs during their burial does seem to provide a satisfactory solution to the problem. -

Ichthyosaur Species Valid Taxa Acamptonectes Fischer Et Al., 2012: Acamptonectes Densus Fischer Et Al., 2012, Lower Cretaceous, Eng- Land, Germany
Ichthyosaur species Valid taxa Acamptonectes Fischer et al., 2012: Acamptonectes densus Fischer et al., 2012, Lower Cretaceous, Eng- land, Germany. Aegirosaurus Bardet and Fernández, 2000: Aegirosaurus leptospondylus (Wagner 1853), Upper Juras- sic–Lower Cretaceous?, Germany, Austria. Arthropterygius Maxwell, 2010: Arthropterygius chrisorum (Russell, 1993), Upper Jurassic, Canada, Ar- gentina?. Athabascasaurus Druckenmiller and Maxwell, 2010: Athabascasaurus bitumineus Druckenmiller and Maxwell, 2010, Lower Cretaceous, Canada. Barracudasauroides Maisch, 2010: Barracudasauroides panxianensis (Jiang et al., 2006), Middle Triassic, China. Besanosaurus Dal Sasso and Pinna, 1996: Besanosaurus leptorhynchus Dal Sasso and Pinna, 1996, Middle Triassic, Italy, Switzerland. Brachypterygius Huene, 1922: Brachypterygius extremus (Boulenger, 1904), Upper Jurassic, Engand; Brachypterygius mordax (McGowan, 1976), Upper Jurassic, England; Brachypterygius pseudoscythius (Efimov, 1998), Upper Jurassic, Russia; Brachypterygius alekseevi (Arkhangelsky, 2001), Upper Jurassic, Russia; Brachypterygius cantabridgiensis (Lydekker, 1888a), Lower Cretaceous, England. Californosaurus Kuhn, 1934: Californosaurus perrini (Merriam, 1902), Upper Triassic USA. Callawayia Maisch and Matzke, 2000: Callawayia neoscapularis (McGowan, 1994), Upper Triassic, Can- ada. Caypullisaurus Fernández, 1997: Caypullisaurus bonapartei Fernández, 1997, Upper Jurassic, Argentina. Chaohusaurus Young and Dong, 1972: Chaohusaurus geishanensis Young and Dong, 1972, Lower Trias- sic, China. -

Geology of the Shepton Mallet Area (Somerset)
Geology of the Shepton Mallet area (Somerset) Integrated Geological Surveys (South) Internal Report IR/03/94 BRITISH GEOLOGICAL SURVEY INTERNAL REPORT IR/03/00 Geology of the Shepton Mallet area (Somerset) C R Bristow and D T Donovan Contributor H C Ivimey-Cook (Jurassic biostratigraphy) The National Grid and other Ordnance Survey data are used with the permission of the Controller of Her Majesty’s Stationery Office. Ordnance Survey licence number GD 272191/1999 Key words Somerset, Jurassic. Subject index Bibliographical reference BRISTOW, C R and DONOVAN, D T. 2003. Geology of the Shepton Mallet area (Somerset). British Geological Survey Internal Report, IR/03/00. 52pp. © NERC 2003 Keyworth, Nottingham British Geological Survey 2003 BRITISH GEOLOGICAL SURVEY The full range of Survey publications is available from the BGS Keyworth, Nottingham NG12 5GG Sales Desks at Nottingham and Edinburgh; see contact details 0115-936 3241 Fax 0115-936 3488 below or shop online at www.thebgs.co.uk e-mail: [email protected] The London Information Office maintains a reference collection www.bgs.ac.uk of BGS publications including maps for consultation. Shop online at: www.thebgs.co.uk The Survey publishes an annual catalogue of its maps and other publications; this catalogue is available from any of the BGS Sales Murchison House, West Mains Road, Edinburgh EH9 3LA Desks. 0131-667 1000 Fax 0131-668 2683 The British Geological Survey carries out the geological survey of e-mail: [email protected] Great Britain and Northern Ireland (the latter as an agency service for the government of Northern Ireland), and of the London Information Office at the Natural History Museum surrounding continental shelf, as well as its basic research (Earth Galleries), Exhibition Road, South Kensington, London projects. -
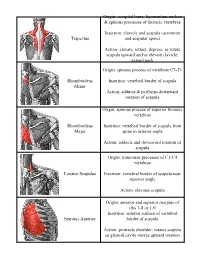
Trapezius Origin: Occipital Bone, Ligamentum Nuchae & Spinous Processes of Thoracic Vertebrae Insertion: Clavicle and Scapul
Origin: occipital bone, ligamentum nuchae & spinous processes of thoracic vertebrae Insertion: clavicle and scapula (acromion Trapezius and scapular spine) Action: elevate, retract, depress, or rotate scapula upward and/or elevate clavicle; extend neck Origin: spinous process of vertebrae C7-T1 Rhomboideus Insertion: vertebral border of scapula Minor Action: adducts & performs downward rotation of scapula Origin: spinous process of superior thoracic vertebrae Rhomboideus Insertion: vertebral border of scapula from Major spine to inferior angle Action: adducts and downward rotation of scapula Origin: transverse precesses of C1-C4 vertebrae Levator Scapulae Insertion: vertebral border of scapula near superior angle Action: elevates scapula Origin: anterior and superior margins of ribs 1-8 or 1-9 Insertion: anterior surface of vertebral Serratus Anterior border of scapula Action: protracts shoulder: rotates scapula so glenoid cavity moves upward rotation Origin: anterior surfaces and superior margins of ribs 3-5 Insertion: coracoid process of scapula Pectoralis Minor Action: depresses & protracts shoulder, rotates scapula (glenoid cavity rotates downward), elevates ribs Origin: supraspinous fossa of scapula Supraspinatus Insertion: greater tuberacle of humerus Action: abduction at the shoulder Origin: infraspinous fossa of scapula Infraspinatus Insertion: greater tubercle of humerus Action: lateral rotation at shoulder Origin: clavicle and scapula (acromion and adjacent scapular spine) Insertion: deltoid tuberosity of humerus Deltoid Action: -

Orbital Pacing and Secular Evolution of the Early Jurassic Carbon Cycle
Orbital pacing and secular evolution of the Early Jurassic carbon cycle Marisa S. Storma,b,1, Stephen P. Hesselboc,d, Hugh C. Jenkynsb, Micha Ruhlb,e, Clemens V. Ullmannc,d, Weimu Xub,f, Melanie J. Lengg,h, James B. Ridingg, and Olga Gorbanenkob aDepartment of Earth Sciences, Stellenbosch University, 7600 Stellenbosch, South Africa; bDepartment of Earth Sciences, University of Oxford, OX1 3AN Oxford, United Kingdom; cCamborne School of Mines, University of Exeter, TR10 9FE Penryn, United Kingdom; dEnvironment and Sustainability Institute, University of Exeter, TR10 9FE Penryn, United Kingdom; eDepartment of Geology, Trinity College Dublin, The University of Dublin, Dublin 2, Ireland; fDepartment of Botany, Trinity College Dublin, The University of Dublin, Dublin 2, Ireland; gEnvironmental Science Centre, British Geological Survey, NG12 5GG Nottingham, United Kingdom; and hSchool of Biosciences, University of Nottingham, LE12 5RD Loughborough, United Kingdom Edited by Lisa Tauxe, University of California San Diego, La Jolla, CA, and approved January 3, 2020 (received for review July 14, 2019) Global perturbations to the Early Jurassic environment (∼201 to where they are recorded as individual shifts or series of shifts ∼174 Ma), notably during the Triassic–Jurassic transition and Toar- within stratigraphically limited sections. Some of these short-term cian Oceanic Anoxic Event, are well studied and largely associated δ13C excursions have been shown to represent changes in the with volcanogenic greenhouse gas emissions released by large supraregional to global carbon cycle, marked by synchronous igneous provinces. The long-term secular evolution, timing, and changes in δ13C in marine and terrestrial organic and inorganic pacing of changes in the Early Jurassic carbon cycle that provide substrates and recorded on a wide geographic extent (e.g., refs. -

Macropredatory Ichthyosaur from the Middle Triassic and the Origin of Modern Trophic Networks
Macropredatory ichthyosaur from the Middle Triassic and the origin of modern trophic networks Nadia B. Fröbischa,1, Jörg Fröbischa,1, P. Martin Sanderb,1,2, Lars Schmitzc,1,2,3, and Olivier Rieppeld aMuseum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung an der Humboldt-Universität zu Berlin, 10115 Berlin, Germany; bSteinmann Institute of Geology, Mineralogy, and Paleontology, Division of Paleontology, University of Bonn, 53115 Bonn, Germany; cDepartment of Evolution and Ecology, University of California, Davis, CA 95616; and dDepartment of Geology, The Field Museum of Natural History, Chicago, IL 60605 Edited by Neil H. Shubin, The University of Chicago, Chicago, IL, and approved December 5, 2012 (received for review October 8, 2012) The biotic recovery from Earth’s most severe extinction event at the Holotype and Only Specimen. The Field Museum of Natural His- Permian-Triassic boundary largely reestablished the preextinction tory (FMNH) contains specimen PR 3032, a partial skeleton structure of marine trophic networks, with marine reptiles assuming including most of the skull (Fig. 1) and axial skeleton, parts of the predator roles. However, the highest trophic level of today’s the pelvic girdle, and parts of the hind fins. marine ecosystems, i.e., macropredatory tetrapods that forage on prey of similar size to their own, was thus far lacking in the Paleozoic Horizon and Locality. FMNH PR 3032 was collected in 2008 from the and early Mesozoic. Here we report a top-tier tetrapod predator, middle Anisian Taylori Zone of the Fossil Hill Member of the Favret a very large (>8.6 m) ichthyosaur from the early Middle Triassic Formation at Favret Canyon, Augusta Mountains, Pershing County, (244 Ma), of Nevada. -

Somerset Geology-A Good Rock Guide
SOMERSET GEOLOGY-A GOOD ROCK GUIDE Hugh Prudden The great unconformity figured by De la Beche WELCOME TO SOMERSET Welcome to green fields, wild flower meadows, farm cider, Cheddar cheese, picturesque villages, wild moorland, peat moors, a spectacular coastline, quiet country lanes…… To which we can add a wealth of geological features. The gorge and caves at Cheddar are well-known. Further east near Frome there are Silurian volcanics, Carboniferous Limestone outcrops, Variscan thrust tectonics, Permo-Triassic conglomerates, sediment-filled fissures, a classic unconformity, Jurassic clays and limestones, Cretaceous Greensand and Chalk topped with Tertiary remnants including sarsen stones-a veritable geological park! Elsewhere in Mendip are reminders of coal and lead mining both in the field and museums. Today the Mendips are a major source of aggregates. The Mesozoic formations curve in an arc through southwest and southeast Somerset creating vales and escarpments that define the landscape and clearly have influenced the patterns of soils, land use and settlement as at Porlock. The church building stones mark the outcrops. Wilder country can be found in the Quantocks, Brendon Hills and Exmoor which are underlain by rocks of Devonian age and within which lie sunken blocks (half-grabens) containing Permo-Triassic sediments. The coastline contains exposures of Devonian sediments and tectonics west of Minehead adjoining the classic exposures of Mesozoic sediments and structural features which extend eastward to the Parrett estuary. The predominance of wave energy from the west and the large tidal range of the Bristol Channel has resulted in rapid cliff erosion and longshore drift to the east where there is a full suite of accretionary landforms: sandy beaches, storm ridges, salt marsh, and sand dunes popular with summer visitors. -

First Amphibian Ichthyosaur Fossil Found
First amphibian ichthyosaur fossil found An ancient marine reptile with seal-like flippers may have been adapted to life on the land as well as in the sea, scientists believe. The 250-million-year-old creature is the first amphibious ichthyosaur known. Its relatives were dolphin-like creatures that swam in the oceans at the time of the dinosaurs. They are thought to have had terrestrial ancestors, but previously no fossils had come to light marking the transition of ichthyosaurs from land to sea. “Now we have this fossil showing the transition,” said lead scientist Professor Ryosuke Motani, from the University of California at Davis who reported the discovery in the journal Nature. At 1.5 feet long, Cartorhynchus lenticarpus was also the smallest known ichthyosaur. Its fossil remains found in Anhui Province, China, date from the start of the Triassic period about 248 million years ago. As well as big flippers, Cartorhynchus had flexible wrists which would have been essential for movement on the ground. While most ichthyosaurs have long beak-like snouts, the new specimen possessed a short nose that may have been adapted to suction feeding. Its body also contained thicker bones than other ichthyosaurs. This supports the theory that most marine reptiles that left the land first grew heavier to help them swim through rough coastal waves. The animal lived about 4 million years after the worst mass extinction in history, shedding light on how long it took for life on Earth to recover. The Permian-Triassic extinction, known as the “Great Dying”, wiped out 96% of all species and may have been linked to global warming. -

Coracoid Process Anatomy: a Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B
Quantitative Assessment of the Coracoacromial and the Coracoclavicular Ligaments With 3-Dimensional Mapping of the Coracoid Process Anatomy: A Cadaveric Study of Surgically Relevant Structures Jorge Chahla, M.D., Ph.D., Daniel Cole Marchetti, B.A., Gilbert Moatshe, M.D., Márcio B. Ferrari, M.D., George Sanchez, B.S., Alex W. Brady, M.Sc., Jonas Pogorzelski, M.D., M.H.B.A., George F. Lebus, M.D., Peter J. Millett, M.D., M.Sc., Robert F. LaPrade, M.D., Ph.D., and CAPT Matthew T. Provencher, M.D., M.C., U.S.N.R. Purpose: To perform a quantitative anatomic evaluation of the (1) coracoid process, specifically the attachment sites of the conjoint tendon, the pectoralis minor, the coracoacromial ligament (CAL), and the coracoclavicular (CC) ligaments in relation to pertinent osseous and soft tissue landmarks; (2) CC ligaments’ attachments on the clavicle; and (3) CAL attachment on the acromion in relation to surgically relevant anatomic landmarks to assist in planning of the Latarjet procedure, acromioclavicular (AC) joint reconstructions, and CAL resection distances avoiding iatrogenic injury to sur- rounding structures. Methods: Ten nonpaired fresh-frozen human cadaveric shoulders (mean age 52 years, range 33- 64 years) were included in this study. A 3-dimensional coordinate measuring device was used to quantify the location of pertinent bony landmarks and soft tissue attachment areas. The ligament and tendon attachment perimeters and center points on the coracoid, clavicle, and acromion were identified and subsequently dissected off the bone. Coordinates of points along the perimeters of attachment sites were used to calculate areas, whereas coordinates of center points were used to determine distances between surgically relevant attachment sites and pertinent bony landmarks. -

Reconstructing Pectoral Appendicular Muscle Anatomy in Fossil Fish and Tetrapods Over the Fins-To-Limbs Transition
Biol. Rev. (2017), pp. 000–000. 1 doi: 10.1111/brv.12386 Reconstructing pectoral appendicular muscle anatomy in fossil fish and tetrapods over the fins-to-limbs transition Julia L. Molnar1,∗ , Rui Diogo2, John R. Hutchinson3 and Stephanie E. Pierce4 1Department of Anatomy, New York Institute of Technology College of Osteopathic Medicine, Northern Boulevard, Old Westbury, NY, U.S.A. 2Department of Anatomy, Howard University College of Medicine, 520 W St. NW, Numa Adams Building, Washington, DC 20059, U.S.A. 3Structure and Motion Lab, Royal Veterinary College, Hawkshead Lane, Hatfield, Hertfordshire AL9 7TA, UK 4Museum of Comparative Zoology and Department of Organismic and Evolutionary Biology, Harvard University, 26 Oxford Street, Cambridge, MA 02138, U.S.A. ABSTRACT The question of how tetrapod limbs evolved from fins is one of the great puzzles of evolutionary biology. While palaeontologists, developmental biologists, and geneticists have made great strides in explaining the origin and early evolution of limb skeletal structures, that of the muscles remains largely unknown. The main reason is the lack of consensus about appendicular muscle homology between the closest living relatives of early tetrapods: lobe-finned fish and crown tetrapods. In the light of a recent study of these homologies, we re-examined osteological correlates of muscle attachment in the pectoral girdle, humerus, radius, and ulna of early tetrapods and their close relatives. Twenty-nine extinct and six extant sarcopterygians were included in a meta-analysis using information from the literature and from original specimens, when possible. We analysed these osteological correlates using parsimony-based character optimization in order to reconstruct muscle anatomy in ancestral lobe-finned fish, tetrapodomorph fish, stem tetrapods, and crown tetrapods.