Myrtaceae, a Cache of Fungal Biodiversity
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Trees for Farm Forestry: 22 Promising Species
Forestry and Forest Products Natural Heritage Trust Helping Communities Helping Australia TREES FOR FARM FORESTRY: 22 PROMISING SPECIES Forestry and Forest Products TREES FOR FARM FORESTRY: Natural Heritage 22 PROMISING SPECIES Trust Helping Communities Helping Australia A report for the RIRDC/ Land & Water Australia/ FWPRDC Joint Venture Agroforestry Program Revised and Edited by Bronwyn Clarke, Ian McLeod and Tim Vercoe March 2009 i © 2008 Rural Industries Research and Development Corporation. All rights reserved. ISBN 1 74151 821 0 ISSN 1440-6845 Trees for Farm Forestry: 22 promising species Publication No. 09/015 Project No. CSF-56A The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable regions. You must not rely on any information contained in this publication without taking specialist advice relevant to your particular circumstances. While reasonable care has been taken in preparing this publication to ensure that information is true and correct, the Commonwealth of Australia gives no assurance as to the accuracy of any information in this publication. The Commonwealth of Australia, the Rural Industries Research and Development Corporation (RIRDC), the authors or contributors expressly disclaim, to the maximum extent permitted by law, all responsibility and liability to any person, arising directly or indirectly from any act or omission, or for any consequences of any such act or omission, made in reliance on the contents of this publication, whether or not caused by any negligence on the part of the Commonwealth of Australia, RIRDC, the authors or contributors. The Commonwealth of Australia does not necessarily endorse the views in this publication. -

Brisbane Native Plants by Suburb
INDEX - BRISBANE SUBURBS SPECIES LIST Acacia Ridge. ...........15 Chelmer ...................14 Hamilton. .................10 Mayne. .................25 Pullenvale............... 22 Toowong ....................46 Albion .......................25 Chermside West .11 Hawthorne................. 7 McDowall. ..............6 Torwood .....................47 Alderley ....................45 Clayfield ..................14 Heathwood.... 34. Meeandah.............. 2 Queensport ............32 Trinder Park ...............32 Algester.................... 15 Coopers Plains........32 Hemmant. .................32 Merthyr .................7 Annerley ...................32 Coorparoo ................3 Hendra. .................10 Middle Park .........19 Rainworth. ..............47 Underwood. ................41 Anstead ....................17 Corinda. ..................14 Herston ....................5 Milton ...................46 Ransome. ................32 Upper Brookfield .......23 Archerfield ...............32 Highgate Hill. ........43 Mitchelton ...........45 Red Hill.................... 43 Upper Mt gravatt. .......15 Ascot. .......................36 Darra .......................33 Hill End ..................45 Moggill. .................20 Richlands ................34 Ashgrove. ................26 Deagon ....................2 Holland Park........... 3 Moorooka. ............32 River Hills................ 19 Virginia ........................31 Aspley ......................31 Doboy ......................2 Morningside. .........3 Robertson ................42 Auchenflower -

Southern Gulf, Queensland
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

BIODIVERSITY CONSERVATION on the TIWI ISLANDS, NORTHERN TERRITORY: Part 1. Environments and Plants
BIODIVERSITY CONSERVATION ON THE TIWI ISLANDS, NORTHERN TERRITORY: Part 1. Environments and plants Report prepared by John Woinarski, Kym Brennan, Ian Cowie, Raelee Kerrigan and Craig Hempel. Darwin, August 2003 Cover photo: Tall forests dominated by Darwin stringybark Eucalyptus tetrodonta, Darwin woollybutt E. miniata and Melville Island Bloodwood Corymbia nesophila are the principal landscape element across the Tiwi islands (photo: Craig Hempel). i SUMMARY The Tiwi Islands comprise two of Australia’s largest offshore islands - Bathurst (with an area of 1693 km 2) and Melville (5788 km 2) Islands. These are Aboriginal lands lying about 20 km to the north of Darwin, Northern Territory. The islands are of generally low relief with relatively simple geological patterning. They have the highest rainfall in the Northern Territory (to about 2000 mm annual average rainfall in the far north-west of Melville and north of Bathurst). The human population of about 2000 people lives mainly in the three towns of Nguiu, Milakapati and Pirlangimpi. Tall forests dominated by Eucalyptus miniata, E. tetrodonta, and Corymbia nesophila cover about 75% of the island area. These include the best developed eucalypt forests in the Northern Territory. The Tiwi Islands also include nearly 1300 rainforest patches, with floristic composition in many of these patches distinct from that of the Northern Territory mainland. Although the total extent of rainforest on the Tiwi Islands is small (around 160 km 2 ), at an NT level this makes up an unusually high proportion of the landscape and comprises between 6 and 15% of the total NT rainforest extent. The Tiwi Islands also include nearly 200 km 2 of “treeless plains”, a vegetation type largely restricted to these islands. -

Guava (Eucalyptus) Rust Puccinia Psidii
INDUSTRY BIOSECURITY PLAN FOR THE NURSERY & GARDEN INDUSTRY Threat Specific Contingency Plan Guava (eucalyptus) rust Puccinia psidii Plant Health Australia March 2009 Disclaimer The scientific and technical content of this document is current to the date published and all efforts were made to obtain relevant and published information on the pest. New information will be included as it becomes available, or when the document is reviewed. The material contained in this publication is produced for general information only. It is not intended as professional advice on any particular matter. No person should act or fail to act on the basis of any material contained in this publication without first obtaining specific, independent professional advice. Plant Health Australia and all persons acting for Plant Health Australia in preparing this publication, expressly disclaim all and any liability to any persons in respect of anything done by any such person in reliance, whether in whole or in part, on this publication. The views expressed in this publication are not necessarily those of Plant Health Australia. Further information For further information regarding this contingency plan, contact Plant Health Australia through the details below. Address: Suite 5, FECCA House 4 Phipps Close DEAKIN ACT 2600 Phone: +61 2 6215 7700 Fax: +61 2 6260 4321 Email: [email protected] Website: www.planthealthaustralia.com.au PHA & NGIA | Contingency Plan – Guava rust (Puccinia psidii) 1 Purpose and background of this contingency plan ............................................................. -

Developing Species for Woody Biomass Crops in Lower Rainfall Southern Australia Australia F FLORASEARCH 3A
Developing Species for Woody Biomass Crops in Lower Rainfall Southern Australia Australia F FLORASEARCH 3A Developing Species for Woody Biomass Crops in Lower Rainfall Southern Australia FloraSearch 3a by Trevor J. Hobbs, Michael Bennell and John Bartle (eds) August 2009 RIRDC Publication No 09/043 RIRDC Project No UWA-98A © 2009 Rural Industries Research and Development Corporation. All rights reserved. ISBN 1 74151 846 6 ISSN 1440-6845 Developing Species for Woody Biomass Crops in Lower Rainfall Southern Australia - FloraSearch 3a Publication No. 09/043 Project No. UWA-98A The information contained in this publication is intended for general use to assist public knowledge and discussion and to help improve the development of sustainable regions. You must not rely on any information contained in this publication without taking specialist advice relevant to your particular circumstances. While reasonable care has been taken in preparing this publication to ensure that information is true and correct, the Commonwealth of Australia gives no assurance as to the accuracy of any information in this publication. The Commonwealth of Australia, the Rural Industries Research and Development Corporation (RIRDC), the authors or contributors expressly disclaim, to the maximum extent permitted by law, all responsibility and liability to any person, arising directly or indirectly from any act or omission, or for any consequences of any such act or omission, made in reliance on the contents of this publication, whether or not caused by any negligence on the part of the Commonwealth of Australia, RIRDC, the authors or contributors. The Commonwealth of Australia does not necessarily endorse the views in this publication. -

Northern Territory NT Page 1 of 204 21-Jan-11 Species List for NRM Region Northern Territory, Northern Territory
Biodiversity Summary for NRM Regions Species List What is the summary for and where does it come from? This list has been produced by the Department of Sustainability, Environment, Water, Population and Communities (SEWPC) for the Natural Resource Management Spatial Information System. The list was produced using the AustralianAustralian Natural Natural Heritage Heritage Assessment Assessment Tool Tool (ANHAT), which analyses data from a range of plant and animal surveys and collections from across Australia to automatically generate a report for each NRM region. Data sources (Appendix 2) include national and state herbaria, museums, state governments, CSIRO, Birds Australia and a range of surveys conducted by or for DEWHA. For each family of plant and animal covered by ANHAT (Appendix 1), this document gives the number of species in the country and how many of them are found in the region. It also identifies species listed as Vulnerable, Critically Endangered, Endangered or Conservation Dependent under the EPBC Act. A biodiversity summary for this region is also available. For more information please see: www.environment.gov.au/heritage/anhat/index.html Limitations • ANHAT currently contains information on the distribution of over 30,000 Australian taxa. This includes all mammals, birds, reptiles, frogs and fish, 137 families of vascular plants (over 15,000 species) and a range of invertebrate groups. Groups notnot yet yet covered covered in inANHAT ANHAT are notnot included included in in the the list. list. • The data used come from authoritative sources, but they are not perfect. All species names have been confirmed as valid species names, but it is not possible to confirm all species locations. -

Bardi Plants an Annotated List of Plants and Their Use
H.,c H'cst. /lust JIus lH8f), 12 (:J): :317-:359 BanE Plants: An Annotated List of Plants and Their Use by the Bardi Aborigines of Dampierland, in North-western Australia \!o\a Smith and .\rpad C. Kalotast Abstract This paper presents a descriptive list of the plants identified and used by the BarcE .\borigines of the Dampierland Peninsula, north~\q:stern Australia. It is not exhaust~ ive. The information is presented in two wavs. First is an alphabetical list of Bardi names including genera and species, use, collection number and references. Second is a list arranged alphabetically according to botanical genera and species, and including family and Bardi name. Previous ethnographic research in the region, vegetation communities and aspects of seasonality (I) and taxonomy arc des~ cribed in the Introduction. Introduction At the time of European colonisation of the south~west Kimberley in the mid nineteenth century, the Bardi Aborigines occupied the northern tip of the Dam pierland Peninsula. To their east lived the island-dwelling Djawi and to the south, the ~yulnyul. Traditionally, Bardi land ownership was based on identification with a particular named huru, translated as home, earth, ground or country. Forty-six bum have been identified (Robinson 1979: 189), and individually they were owned by members of a family tracing their ownership patrilineally, and known by the bum name. Collectively, the buru fall into four regions with names which are roughly equivalent to directions: South: Olonggong; North-west: Culargon; ~orth: Adiol and East: Baniol (Figure 1). These four directional terms bear a superficial resemblance to mainland subsection kinship patterns, in that people sometimes refer to themselves according to the direction in which their land lies, and indeed 'there are. -

Accuracy Assessment of Vegetation Community Maps Generated by Aerial Photography Interpretation: Perspective from the Tropical Savanna, Australia
Accuracy assessment of vegetation community maps generated by aerial photography interpretation: perspective from the tropical savanna, Australia Donna L. Lewis Stuart Phinn Downloaded From: http://remotesensing.spiedigitallibrary.org/ on 10/13/2015 Terms of Use: http://spiedigitallibrary.org/ss/TermsOfUse.aspx Accuracy assessment of vegetation community maps generated by aerial photography interpretation: perspective from the tropical savanna, Australia Donna L. Lewisa,b and Stuart Phinna a The University of Queensland, School of Geography, Planning and Environmental Management, Centre for Spatial Environmental Research, Biophysical Remote Sensing Group, Brisbane, Queensland, Australia, 4072 [email protected]; [email protected] b Department of Natural Resources, Environment, The Arts and Sport, P.O. Box 496, Palmerston, NT 0831, Australia [email protected] Abstract. Aerial photography interpretation is the most common mapping technique in the world. However, unlike an algorithm-based classification of satellite imagery, accuracy of aerial photography interpretation generated maps is rarely assessed. Vegetation communities covering an area of 530 km2 on Bullo River Station, Northern Territory, Australia, were mapped using an interpretation of 1:50,000 color aerial photography. Manual stereoscopic line-work was delineated at 1:10,000 and thematic maps generated at 1:25,000 and 1:100,000. Multi- variate and intuitive analysis techniques were employed to identify 22 vegetation communities within the study area. The accuracy assessment was based on 50% of a field dataset collected over a 4 year period (2006 to 2009) and the remaining 50% of sites were used for map attri- bution. The overall accuracy and Kappa coefficient for both thematic maps was 66.67% and 0.63, respectively, calculated from standard error matrices. -

Defining and Mapping Rare Vegetation Communities: Improving Techniques to Assist Land-Use Planning and Conservation
Defining and mapping rare vegetation communities: improving techniques to assist land-use planning and conservation Stephen A. J. Bell BSc. (Hons) A thesis submitted for the degree of Doctor of Philosophy School of Environmental and Life Sciences The University of Newcastle February 2013 Declaration Statement of Originality This thesis contains no material which has been accepted for the award of any other degree or diploma in any university or other tertiary institution and, to the best of my knowledge and belief, contains no material previously published or written by another person, except where due reference has been made in the text. I give consent to this copy of my thesis, when deposited in the University Library, being made available for loan and photocopying subject to the provisions of the Copyright Act 1968. Statement of Collaboration I hereby certify that the concepts embodied in Chapter 2 of this thesis have been done in collaboration with a fellow researcher at this university. I have included as part of the thesis in Chapter 2 a statement clearly outlining the extent of collaboration with whom and under what auspices. Stephen A. J. Bell Preface “The vegetated landscape …. on first appearance presents a bewildering display of living matter, a higgledy-piggledy mass of trunks, leaves, branches, shrubs and grasses seemingly without form. The more observant may notice that the higgledy-piggledy mass varies from one place to another, that in some places there are trees as tall as large buildings while in other places there are no trees at all …. By the application of a systematic approach to viewing vegetation the bewildering display of plant life can take on new meaning thus altering one’s perception of what is being seen .… Suddenly the jumble of plant life reveals structures and beauties probably hitherto unseen”. -
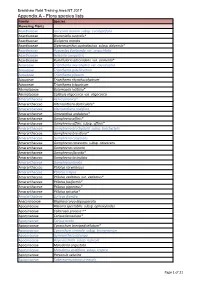
Report-NT-Bradshaw-Appendix A
Bradshaw Field Training Area NT 2017 Appendix A - Flora species lists Family Species Flowering Plants Acanthaceae Avicennia marina subsp. eucalyptifolia Acanthaceae Brunoniella australis* Acanthaceae Dicliptera armata Acanthaceae Dipteracanthus australasicus subsp. dalyensis* Acanthaceae Hypoestes floribunda var. angustifolia Acanthaceae Nelsonia campestris Acanthaceae Rostellularia adscendens var. clementii* Aizoaceae Trianthema oxycalyptra var. oxycalyptra Aizoaceae Trianthema patellitectum Aizoaceae Trianthema pilosum Aizoaceae Trianthema rhynchocalyptrum Aizoaceae Trianthema triquetrum Alismataceae Butomopsis latifolia* Alismataceae Caldesia oligococca var. oligococca Amaranthaceae Aerva javanica^ Amaranthaceae Alternanthera denticulata* Amaranthaceae Alternanthera nodiflora Amaranthaceae Amaranthus undulatus* Amaranthaceae Gomphrena affinis* Amaranthaceae Gomphrena affinis subsp. affinis* Amaranthaceae Gomphrena brachystylis subsp. brachystylis Amaranthaceae Gomphrena breviflora* Amaranthaceae Gomphrena canescens Amaranthaceae Gomphrena canescens subsp. canescens Amaranthaceae Gomphrena connata Amaranthaceae Gomphrena flaccida* Amaranthaceae Gomphrena lacinulata Amaranthaceae Gomphrena lanata Amaranthaceae Ptilotus corymbosus Amaranthaceae Ptilotus crispus Amaranthaceae Ptilotus exaltatus var. exaltatus* Amaranthaceae Ptilotus fusiformis* Amaranthaceae Ptilotus giganteus* Amaranthaceae Ptilotus spicatus* Amaranthaceae Surreya diandra Anacardiaceae Blepharocarya depauperata Apocynaceae Alstonia spectabilis subsp. ophioxyloides Apocynaceae -

Descriptions of the Plant Types
APPENDIX A Descriptions of the plant types The plant life forms employed in the model are listed, with examples, in the main text (Table 2). They are described in this appendix in more detail, including environmental relations, physiognomic characters, prototypic and other characteristic taxa, and relevant literature. A list of the forms, with physiognomic characters, is included. Sources of vegetation data relevant to particular life forms are cited with the respective forms in the text of the appendix. General references, especially descriptions of regional vegetation, are listed by region at the end of the appendix. Plant form Plant size Leaf size Leaf (Stem) structure Trees (Broad-leaved) Evergreen I. Tropical Rainforest Trees (lowland. montane) tall, med. large-med. cor. 2. Tropical Evergreen Microphyll Trees medium small cor. 3. Tropical Evergreen Sclerophyll Trees med.-tall medium seier. 4. Temperate Broad-Evergreen Trees a. Warm-Temperate Evergreen med.-small med.-small seier. b. Mediterranean Evergreen med.-small small seier. c. Temperate Broad-Leaved Rainforest medium med.-Iarge scler. Deciduous 5. Raingreen Broad-Leaved Trees a. Monsoon mesomorphic (lowland. montane) medium med.-small mal. b. Woodland xeromorphic small-med. small mal. 6. Summergreen Broad-Leaved Trees a. typical-temperate mesophyllous medium medium mal. b. cool-summer microphyllous medium small mal. Trees (Narrow and needle-leaved) Evergreen 7. Tropical Linear-Leaved Trees tall-med. large cor. 8. Tropical Xeric Needle-Trees medium small-dwarf cor.-scler. 9. Temperate Rainforest Needle-Trees tall large-med. cor. 10. Temperate Needle-Leaved Trees a. Heliophilic Large-Needled medium large cor. b. Mediterranean med.-tall med.-dwarf cor.-scler.