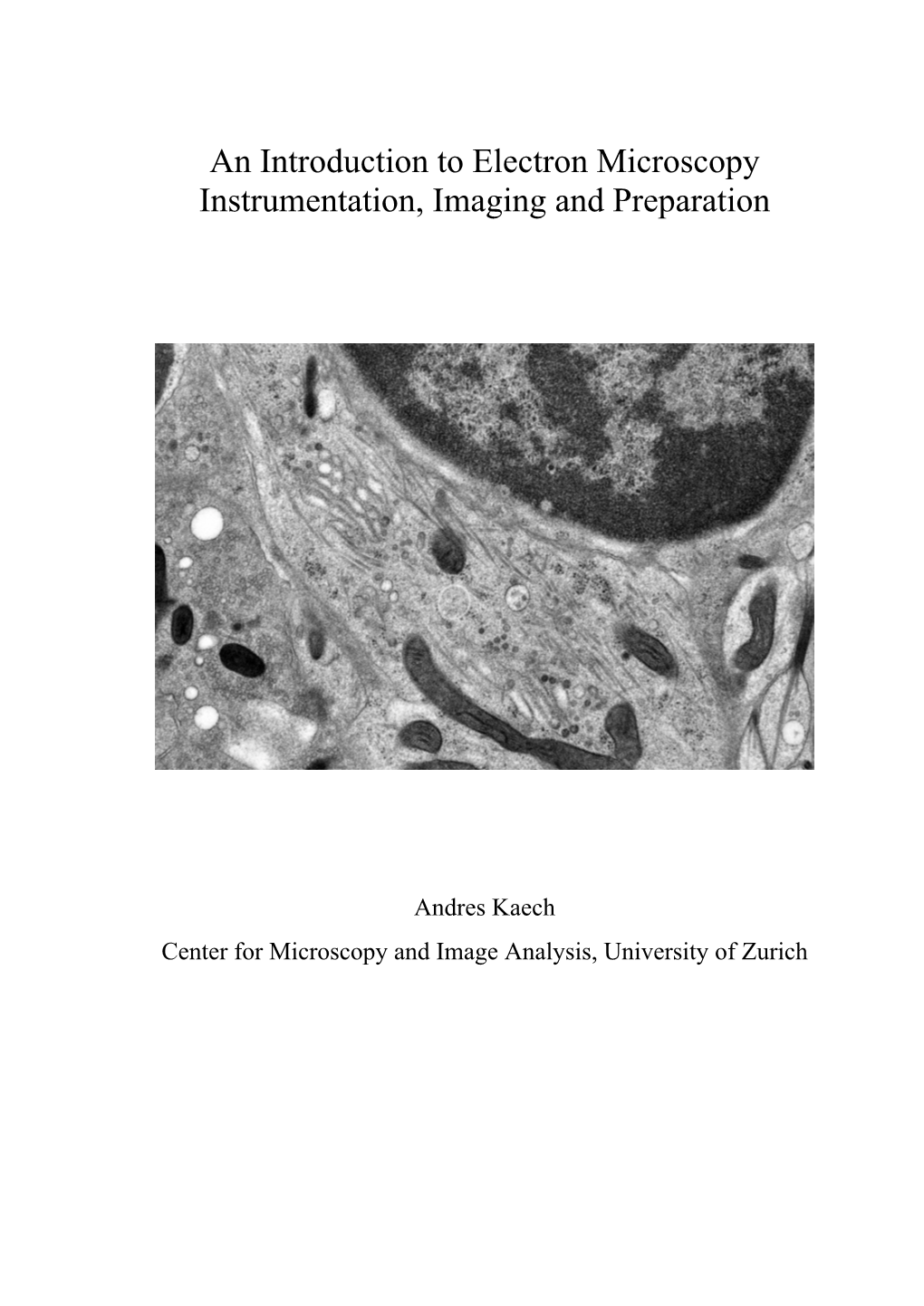
An Introduction to Electron Microscopy Instrumentation, Imaging and Preparation
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Recent Progress in Scanning Electron Microscopy for the Characterization of fine Structural Details of Nano Materials
Progress in Solid State Chemistry 42 (2014) 1e21 Contents lists available at ScienceDirect Progress in Solid State Chemistry journal homepage: www.elsevier.com/locate/pssc Recent progress in scanning electron microscopy for the characterization of fine structural details of nano materials Mitsuo Suga a,*, Shunsuke Asahina a, Yusuke Sakuda a, Hiroyoshi Kazumori a, Hidetoshi Nishiyama a, Takeshi Nokuo a, Viveka Alfredsson b, Tomas Kjellman b, Sam M. Stevens c, Hae Sung Cho d, Minhyung Cho d, Lu Han e, Shunai Che e, Michael W. Anderson f, Ferdi Schüth g, Hexiang Deng h, Omar M. Yaghi i, Zheng Liu j, Hu Young Jeong k, Andreas Stein l, Kazuyuki Sakamoto m, Ryong Ryoo n,o, Osamu Terasaki d,p,** a JEOL Ltd., SM Business Unit, Tokyo, Japan b Physical Chemistry, Lund University, Sweden c Private Contributor, UK d Graduate School of EEWS, KAIST, Republic of Korea e School of Chemistry & Chemical Engineering, Shanghai Jiao Tong University, China f Centre for Nanoporous Materials, School of Chemistry, University of Manchester, UK g Department of Heterogeneous Catalysis, Max-Planck-Institut für Kohlenforschung, Mülheim, Germany h College of Chemistry and Molecular Sciences, Wuhan University, China i Department of Chemistry, University of California, Berkeley, USA j Nanotube Research Center, AIST, Tsukuba, Japan k UNIST Central Research Facilities/School of Mechanical & Advanced Materials Engineering, UNIST, Republic of Korea l Department of Chemistry, University of Minnesota, Minneapolis, USA m Department of Nanomaterials Science, Chiba University, -

Transmission Electron Microscope
GG 711: Advanced Techniques in Geophysics and Materials Science Lecture 14 Transmission Electron Microscope Pavel Zinin HIGP, University of Hawaii, Honolulu, USA www.soest.hawaii.edu\~zinin Resolution of SEM Transmission electron microscopy (TEM) is a microscopy technique whereby a beam of electrons is transmitted through an ultra thin specimen, interacting with the specimen as it passes through. An image is formed from the interaction of the electrons transmitted through the specimen; the image is magnified and focused onto an imaging device, such as a fluorescent screen, on a layer of photographic film, or to be detected by a sensor such as a CCD camera. The first TEM was built by Max Kroll and Ernst Ruska in 1931, with this group developing the first TEM with resolution power greater than that of light in 1933 and the first commercial TEM in 1939. The first practical TEM, Originally installed at I. G Farben-Werke and now on display at the Deutsches Museum in Munich, Germany Comparison of LM and TEM Both glass and EM lenses subject to same distortions and aberrations Glass lenses have fixed focal length, it requires to change objective lens to change magnification. We move objective lens closer to or farther away from specimen to focus. EM lenses to specimen distance fixed, focal length varied by varying current through lens LM: (a) Direct observation TEM: (a) Video imaging (CRT); (b) image of the image; (b) image is is formed by transmitted electrons impinging formed by transmitted light on phosphor coated screen Transmission Electron Microscope: Principle Ray diagram of a conventional transmission electron microscope (top Positioning of signal path) and of a scanning transmission electron microscope (bottom path). -

Light and Electron Microscopy of the Exocrine Pancreas in the Chronically Reserpinized Rat1
003 1 -3998/89/2505-0482$02.00/0 PEDIATRIC RESEARCH Vol. 25, No. 5, 1989 Copyright O 1989 International Pediatric Research Foundation, Inc. Printed rn U.S.A. Light and Electron Microscopy of the Exocrine Pancreas in the Chronically Reserpinized Rat1 GILLES GRONDIN, FRANCOIS A. LEBLOND, JEAN MORISSET, AND DENIS LEBEL Centre de Recherche sur Ies Micanismes de Sicrition, Faculty of Science, University of Sherbrooke, Sherbrooke, QC, Canada, JIK 2RI ABSTRACT. The effects of reserpine injections were stud- been the most studied (1-8). The main effects induced by the ied on the morphology of the pancreas in an experimental reserpine treatment on the ultrastructure of the pancreas are the model for cystic fibrosis, the chronically reserpinized rat. following: an accumulation of granules in the acinar cell, an A detailed examination of the tissue was carried out at the alteration of the granule ultrastructure, a slight diminution of light and electron microscopic levels. The nonspecific ef- the RER and Golgi apparatus, the presence of autophagic bodies, fects of secondary malnutrition induced by the drug were and an augmentation of lysosomal bodies (3,5,9, 10). In another assessed with a group of animals pair fed with the treated paper (1 I), we report the effects of such a treatment on pancreatic animals. In a companion paper, we show that pancreatic growth and on the amount of a glycoprotein characteristic of the wt, lipase, and GP-2 contents also are affected by reserpine zymogen granule membrane, the GP-2. In this study, we specif- treatment. In this study, we report that no morphologic ically report the effects of chronic reserpine treatment at two differences were observed between the exocrine pancreatic doses, on the structure and ultrastructure of the rat pancreas. -

The Future of Electron Microscopy
BNL-108108-2015-JA Feature Article for Physics Today The Future of Electron Microscopy Yimei Zhu and Hermann Dürr Seeing is believing. So goes the old adage, and seen evidence is undoubtedly satisfying in that it can be interpreted readily, though not necessarily always correctly. For centuries, humans have tried continuously to improve their abilities of seeing things, from developing telescopes to observe our solar systems, to microscopes to reveal bacteria and viruses. The 2014 Nobel Prize in Chemistry was awarded to Eric Betzig, Stefan Hell and William Moerner for their pioneering work on super-resolution fluorescence microscopy, overcoming Abbe’s diffraction limit in the resolution of conventional light microscopes. It was a breakthrough that enabled ground-breaking discoveries in biological research and also is testimony of the importance of modern microscopy. In February 2014 the US Department of Energy’s Office of Basic Energy Science held a two day workshop that brought together experts from various fields to identify new science that will be enabled by advances in electron microscopy. [1] The workshop focused on studies of ultrafast processes, atomic resolution, sample environments and measuring functionality at nanoscales. This article captures some of the exciting discussions the workshop generated. Seeing with electrons Electrons, photons and neutrons are three fundamental probes of condensed matter. They are routinely used to study complementary physical properties of materials. Neutrons interact with nuclei and atomic spins, x-ray photons with electron clouds and electron probes with electrostatic potentials, i.e., positively charged nuclei screened by negatively charged electrons. Compared with x-rays and neutrons, one advantage of electrons is their much stronger interaction with matter (105 times stronger than elastically scattered x-rays due to the coulomb interaction); hence, for a very small probing volume, the interaction generates strong signals, making it far easier to image and detect individual atoms, molecules, and nanoscale objects. -

Super-Resolution Imaging by Dielectric Superlenses: Tio2 Metamaterial Superlens Versus Batio3 Superlens
hv photonics Article Super-Resolution Imaging by Dielectric Superlenses: TiO2 Metamaterial Superlens versus BaTiO3 Superlens Rakesh Dhama, Bing Yan, Cristiano Palego and Zengbo Wang * School of Computer Science and Electronic Engineering, Bangor University, Bangor LL57 1UT, UK; [email protected] (R.D.); [email protected] (B.Y.); [email protected] (C.P.) * Correspondence: [email protected] Abstract: All-dielectric superlens made from micro and nano particles has emerged as a simple yet effective solution to label-free, super-resolution imaging. High-index BaTiO3 Glass (BTG) mi- crospheres are among the most widely used dielectric superlenses today but could potentially be replaced by a new class of TiO2 metamaterial (meta-TiO2) superlens made of TiO2 nanoparticles. In this work, we designed and fabricated TiO2 metamaterial superlens in full-sphere shape for the first time, which resembles BTG microsphere in terms of the physical shape, size, and effective refractive index. Super-resolution imaging performances were compared using the same sample, lighting, and imaging settings. The results show that TiO2 meta-superlens performs consistently better over BTG superlens in terms of imaging contrast, clarity, field of view, and resolution, which was further supported by theoretical simulation. This opens new possibilities in developing more powerful, robust, and reliable super-resolution lens and imaging systems. Keywords: super-resolution imaging; dielectric superlens; label-free imaging; titanium dioxide Citation: Dhama, R.; Yan, B.; Palego, 1. Introduction C.; Wang, Z. Super-Resolution The optical microscope is the most common imaging tool known for its simple de- Imaging by Dielectric Superlenses: sign, low cost, and great flexibility. -

Development of Optical Hyperlens for Imaging Below the Diffraction Limit
Development of optical hyperlens for imaging below the diffraction limit Hyesog Lee, Zhaowei Liu, Yi Xiong, Cheng Sun and Xiang Zhang* 5130 Etcheverry Hall, NSF Nanoscale Science and Engineering Center (NSEC), University of California, Berkeley, CA 94720 *Corresponding author: [email protected] http://xlab.me.berkeley.edu Abstract: We report here the design, fabrication and characterization of optical hyperlens that can image sub-diffraction-limited objects in the far field. The hyperlens is based on an artificial anisotropic metamaterial with carefully designed hyperbolic dispersion. We successfully designed and fabricated such a metamaterial hyperlens composed of curved silver/alumina multilayers. Experimental results demonstrate far-field imaging with resolution down to 125nm at 365nm working wavelength which is below the diffraction limit. ©2007 Optical Society of America OCIS codes: (110 0180) Microscopy; (220.4241) Optical design and fabrication: Nanostructure fabrication References and links 1. E. Abbe, Arch. Mikroskop. Anat. 9, 413 (1873) 2. E. Betzig, J. K. Trautman, T. D. Harris, J. S. Weiner and R. L. Kostelak, “Breaking the diffraction barrier – optical microscopy on a nanometric scale,” Science 251, 1468-1470 (1991) 3. S. W. Hell, “Toward Fluorescence nanoscopy,” Nat. Biotechnol. 21, 1347-1355 (2003) 4. M. G. L. Gustafsson, “Nonlinear structured-illumination microscopy: Wide-field fluorescence imaging with theoretically unlimited resolution,” P. Natl. Acad. Sci. 102, 13081-13086 (2005) 5. J. B. Pendry, “Negative refraction makes a perfect lens,” Phys. Rev. Lett. 85, 3966-3969 (2000) 6. N. Fang, H. Lee, C. Sun and X. Zhang, “Sub-Diffraction-Limited Optical Imaging with a Silver Superlens” Science 308, 534-537 (2005) 7. -

Crystallography Beyond Crystals: PX and Spcryoem
GENERAL ARTICLE Crystallography beyond Crystals: PX and SPCryoEM Ramanathan Natesh Protein Crystallography (PX) or Macromolecular Crystal- lography has been unarguably one of the great intellectual achievements of 20th century. In fact, it has developed to be a strong base for ‘hybrid methods’ for macromolecular struc- ture determination by a combination of PX and Single Par- ticle Cryo Electron Microscopy (SPCryoEM). This article describes how PX and SPCryoEM are independent methods, how they contribute as hybrid methods, their complementarity Ramanathan Natesh is a Ramalingaswami Fellow – and supplementarity and their current status. This article DBT and Assistant Professor though not a complete review of PX or SPCryoEM of macro- at the School of Biology, molecular protein complexes, serves as a guideline in under- Indian Institute of Science standing the underlying principles. Education and Research, Thiruvananthapuram. His research interests involve 1. Introduction Molecular Structural Biology studies of proteins Despite the knowledge and popular use of PX techniques in the and its complexes in human scientific community, it is less known to the general public. The health and diseases using International Year of Crystallography (IYCr) 2014 is being orga- two principle techniques viz., Protein Crystallogra- nized jointly by the International Union of Crystallography (IUCr) phy and Single Particle and UNESCO, which has taken many measures to popularize Cryo Electron Microscopy crystallography. A UNESCO brochure on Crystallography Mat- along with range of other ters can be found here http://www.iycr2014.org/about/promo- biophysical and biochemical techniques. tional-materials. One question you might ask is, ‘why is knowing the protein 1 Inhibitors are also called as structure important?’ All living beings are made of cells and there drug molecules that stop the are thousands of proteins that function within and outside these function of the protein by bind- cells. -

Introduction to Electron Microscopy
Introduction to Electron Microscopy Light microscopes were developed in the early 1600’s. By the end of the century Dutch microscopist van Leeuwenhoek had imaged blood cells, bacteria and structure within the cell using a simple one-lens microscope with x300 magnification which was awkward to use. At the same time the compound microscope was being developed. This uses at least two lenses; an objective, placed close to the specimen, and an eyepiece (ocular), placed Diagram of a typical three lens compound microscope close to the eye. Such was van with transmission illumination. Leeuwenhoek’s skill, however, that it was two centuries before the compound microscope could match his work. Compound optical microscopes, these days, have a maximum magnification of about x1000 and an imaging resolution of around 0.3µm for light in the middle of the visible region (λ~0.5µm). Aberrations in light optical lenses can be reduced to a minimum by grinding the lens surface to a correct shape or spacing the lenses so that their aberrations are compensated. In the absence of aberrations the limit of resolution (d) of any wave optical microscope is given by the Rayleigh criterion:- d = 0.61 λ/(µ sin α) where λ is the wavelength of the light (or electrons), µ is the refractive index and α the illumination semi-angle. To get any improvement in resolution either λ needs to be reduced (ultraviolet region), or µ increased (oil immersion). There are two basic forms of light optical microscopes. For specimens that are optically transparent the light can pass from the source, through the specimen, and into the microscope (transmission). -

Molecular Scale Imaging with a Smooth Superlens
Molecular Scale Imaging with a Smooth Superlens Pratik Chaturvedi1, Wei Wu2, VJ Logeeswaran3, Zhaoning Yu2, M. Saif Islam3, S.Y. Wang2, R. Stanley Williams2, & Nicholas Fang1* 1Department of Mechanical Science & Engineering, University of Illinois at Urbana- Champaign, 1206 W. Green St., Urbana, IL 61801, USA. 2Information & Quantum Systems Lab, Hewlett-Packard Laboratories, 1501 Page Mill Rd, MS 1123, Palo Alto, CA 94304, USA. 3Department of Electrical & Computer Engineering, Kemper Hall, University of California at Davis, One Shields Ave, Davis, CA 95616, USA. * Corresponding author Email: [email protected] RECEIVED DATE Abstract We demonstrate a smooth and low loss silver (Ag) optical superlens capable of resolving features at 1/12th of the illumination wavelength with high fidelity. This is made possible by utilizing state-of-the-art nanoimprint technology and intermediate 1 wetting layer of germanium (Ge) for the growth of flat silver films with surface roughness at sub-nanometer scales. Our measurement of the resolved lines of 30nm half-pitch shows a full-width at half-maximum better than 37nm, in excellent agreement with theoretical predictions. The development of this unique optical superlens lead promise to parallel imaging and nanofabrication in a single snapshot, a feat that are not yet available with other nanoscale imaging techniques such as atomic force microscope or scanning electron microscope. λ = 380nm 250nm The resolution of optical images has historically been constrained by the wavelength of light, a well known physical law which is termed as the diffraction limit. Conventional optical imaging is only capable of focusing the propagating components from the source. The evanescent components which carry the subwavelength information exponentially decay in a medium with positive permittivity (ε), and positive permeability (µ) and hence, are lost before making it to the image plane. -

Micro Beams in Physical and Chemical Analytical Applications
Micro Beams in Physical and Chemical Analytical Applications Stjepko Fazinić Laboratory for Ion Beam Interactions Rudjer Bošković Institute, Zagreb, Croatia International Topical Meeting on Nuclear Research Applications and Utilization of Accelerators , Vienna, 4-8 May 2009 Satellite meeting II: Particle Accelerators in Analytical and Educational Applications Ru đer Boškovi ć Institute, Zagreb, Croatia Terminology/instrumentation Particle accelerators and Micro Beams Micro Beams and Microprobes: A microprobe is an instrument that applies a stable and well-focused (micro!) beam of charged particles (electrons or ions) to a sample Focused Ion Beams (FIB) instruments electron microprobes (like scanning electron microscope) ion microprobe: two different clases of instruments employing SIMS (Secondary Ion Mass Spectrometry) nuclear microprobe (nuclear microscope) Another Micro Beams associated with accelerators: accelerator based (mainly synchrotron) x-ray Micro Beams Focused Ion Beams (FIB) usually gallium ions accelerated to 5-50 keV focused onto the sample by electrostatic lenses site-specific analysis, deposition and ablation of materials micro-machining tool (sputtering) ion beam induced deposition of material modifying existing semiconductor device sample preparation for TEM variation: helium ion microscope Helium ions with 25-30 keV energy for surface imaging and material surface composition analysis by Rutherford Backscatering (RBS). Spatial resolution less than 0.9 nm high quality images: comparable or better than -

Macromolecular Mass Spectrometry and Electron Microscopy As Complementary Tools for Investigation of the Heterogeneity of Bacteriophage Portal Assemblies
Journal of Structural Biology Journal of Structural Biology 157 (2007) 371–383 www.elsevier.com/locate/yjsbi Macromolecular mass spectrometry and electron microscopy as complementary tools for investigation of the heterogeneity of bacteriophage portal assemblies Anton Poliakov a,1, Esther van Duijn b,1, Gabriel Lander c, Chi-yu Fu b, John E. Johnson c, Peter E. Prevelige Jr. a,*, Albert J.R. Heck b,* a Department of Microbiology, University of Alabama at Birmingham, Birmingham, AL 35294, USA b Department of Biomolecular Mass Spectrometry, Bijvoet Center for Biomolecular Research and Utrecht Institute for Pharmaceutical Sciences, Utrecht University, Sorbonnelaan 16, 3584 CA Utrecht, The Netherlands c Department of Molecular Biology, The Scripps Research Institute, 10550 North Torrey Pines Road, La Jolla, CA 92037, USA Received 19 July 2006; received in revised form 8 September 2006; accepted 8 September 2006 Available online 19 September 2006 Abstract The success of electron-cryo microscopy (cryo-EM) and image reconstruction of cyclic oligomers, such as the viral and bacteriophage portals, depends on the accurate knowledge of their order of symmetry. A number of statistical methods of image analysis address this problem, but often do not provide unambiguous results. Direct measurement of the oligomeric state of multisubunit protein assemblies is difficult when the number of subunits is large and one subunit renders only a small increment to the full size of the oligomer. Moreover, when mixtures of different stochiometries are present techniques such as analytical centrifugation or size-exclusion chromatography are also less helpful. Here, we use electrospray ionization mass spectrometry to directly determine the oligomeric states of the in vitro assem- bled portal oligomers of the phages P22, Phi-29 and SPP1, which range in mass from 430 kDa to about 1 million Da. -

Current Status and Future Directions for in Situ Transmission Electron Microscopy
1 Current status and future directions for in situ transmission electron microscopy Mitra L. Taheri1, Eric A. Stach2, Ilke Arslan3, P.A. Crozier4, Bernd C. Kabius5,, Thomas LaGrange6,**, Andrew M. Minor7, Seiji Takeda8, Mihaela Tanase9,#, Jakob B. Wagner10, and Renu Sharma9,! 1Department of Materials Science and Engineering, Drexel University 2Center for Functional Nanomaterials, Brookhaven, National Laboratory 3Pacific Northwest National Laboratory, Physical and Computational Sciences Directorate, 902 Battelle Blvd, Richland WA, USA 4School for Engineering of Matter, Transport and Energy, Arizona State University, Tempe, AZ 85281 (USA) 5The Pennsylvania State University, University Park, PA 16802 (USA) 6Lawrence Livermore National Laboratory, Physical and Life Science Directorate, Condensed Matter and Materials Division, 7000 East Avenue, P.O. 808 L-356 7Department of Materials Science & Engineering, University of California, Berkeley and National Center for Electron Microscopy, Molecular Foundry, Lawrence Berkeley National Laboratory, One Cyclotron Road, MS 72, Berkeley, CA USA 8Institute of Scientific and Industrial Research (ISIR), Osaka University, 8-1 Mihogaoka, Ibaraki, Osaka 567-0047, Japan 9 Center for Nanoscale Science and Technology- National Institute of Standards and Technology, Gaithersburg, MD 20899-6203. 10Center for Electron Nanoscopy, Technical University of Denmark, Kgs. Lyngby, Denmark ** Current Address: École Polytechnique Fédérale de Lausanne, Interdisciplinary Centre for Electron Microscopy, EPFL-SB-CIME-GE, MXC