A Review of Avian Influenza a Virus Associations in Synanthropic Birds
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

156 Glossy Ibis
Text and images extracted from Marchant, S. & Higgins, P.J. (co-ordinating editors) 1990. Handbook of Australian, New Zealand & Antarctic Birds. Volume 1, Ratites to ducks; Part B, Australian pelican to ducks. Melbourne, Oxford University Press. Pages 953, 1071-1 078; plate 78. Reproduced with the permission of Bird life Australia and Jeff Davies. 953 Order CICONIIFORMES Medium-sized to huge, long-legged wading birds with well developed hallux or hind toe, and large bill. Variations in shape of bill used for recognition of sub-families. Despite long legs, walk rather than run and escape by flying. Five families of which three (Ardeidae, Ciconiidae, Threskiornithidae) represented in our region; others - Balaenicipitidae (Shoe-billed Stork) and Scopidae (Hammerhead) - monotypic and exclusively Ethiopian. Re lated to Phoenicopteriformes, which sometimes considered as belonging to same order, and, more distantly, to Anseriformes. Behavioural similarities suggest affinities also to Pelecaniformes (van Tets 1965; Meyerriecks 1966), but close relationship not supported by studies of egg-white proteins (Sibley & Ahlquist 1972). Suggested also, mainly on osteological and other anatomical characters, that Ardeidae should be placed in separate order from Ciconiidae and that Cathartidae (New World vultures) should be placed in same order as latter (Ligon 1967). REFERENCES Ligon, J.D. 1967. Occas. Pap. Mus. Zool. Univ. Mich. 651. Sibley, C. G., & J.E. Ahlquist. 1972. Bull. Peabody Mus. nat. Meyerriecks, A.J. 1966. Auk 83: 683-4. Hist. 39. van Tets, G.F. 1965. AOU orn. Monogr. 2. 1071 Family PLATALEIDAE ibises, spoonbills Medium-sized to large wading and terrestial birds. About 30 species in about 15 genera, divided into two sub families: ibises (Threskiornithinae) and spoonbills (Plataleinae); five species in three genera breeding in our region. -

January 2020.Indd
BROWN PELICAN Photo by Rob Swindell at Melbourne, Florida JANUARY 2020 Editors: Jim Jablonski, Marty Ackermann, Tammy Martin, Cathy Priebe Webmistress: Arlene Lengyel January 2020 Program Tuesday, January 7, 2020, 7 p.m. Carlisle Reservation Visitor Center Gulls 101 Chuck Slusarczyk, Jr. "I'm happy to be presenting my program Gulls 101 to the good people of Black River Audubon. Gulls are notoriously difficult to identify, but I hope to at least get you looking at them a little closer. Even though I know a bit about them, I'm far from an expert in the field and there is always more to learn. The challenge is to know the particular field marks that are most important, and familiarization with the many plumage cycles helps a lot too. No one will come out of this presentation an expert, but I hope that I can at least give you an idea what to look for. At the very least, I hope you enjoy the photos. Looking forward to seeing everyone there!” Chuck Slusarczyk is an avid member of the Ohio birding community, and his efforts to assist and educate novice birders via social media are well known, yet he is the first to admit that one never stops learning. He has presented a number of programs to Black River Audubon, always drawing a large, appreciative gathering. 2019 Wellington Area Christmas Bird Count The Wellington-area CBC will take place Saturday, December 28, 2019. Meet at the McDonald’s on Rt. 58 at 8:00 a.m. The leader is Paul Sherwood. -

Influenza Virus Infections in Humans October 2018
Influenza virus infections in humans October 2018 This note is provided in order to clarify the differences among seasonal influenza, pandemic influenza, and zoonotic or variant influenza. Seasonal influenza Seasonal influenza viruses circulate and cause disease in humans every year. In temperate climates, disease tends to occur seasonally in the winter months, spreading from person-to- person through sneezing, coughing, or touching contaminated surfaces. Seasonal influenza viruses can cause mild to severe illness and even death, particularly in some high-risk individuals. Persons at increased risk for severe disease include pregnant women, the very young and very old, immune-compromised people, and people with chronic underlying medical conditions. Seasonal influenza viruses evolve continuously, which means that people can get infected multiple times throughout their lives. Therefore the components of seasonal influenza vaccines are reviewed frequently (currently biannually) and updated periodically to ensure continued effectiveness of the vaccines. There are three large groupings or types of seasonal influenza viruses, labeled A, B, and C. Type A influenza viruses are further divided into subtypes according to the specific variety and combinations of two proteins that occur on the surface of the virus, the hemagglutinin or “H” protein and the neuraminidase or “N” protein. Currently, influenza A(H1N1) and A(H3N2) are the circulating seasonal influenza A virus subtypes. This seasonal A(H1N1) virus is the same virus that caused the 2009 influenza pandemic, as it is now circulating seasonally. In addition, there are two type B viruses that are also circulating as seasonal influenza viruses, which are named after the areas where they were first identified, Victoria lineage and Yamagata lineage. -

Bird Vulnerability Assessments
Assessing the vulnerability of native vertebrate fauna under climate change, to inform wetland and floodplain management of the River Murray in South Australia: Bird Vulnerability Assessments Attachment (2) to the Final Report June 2011 Citation: Gonzalez, D., Scott, A. & Miles, M. (2011) Bird vulnerability assessments- Attachment (2) to ‘Assessing the vulnerability of native vertebrate fauna under climate change to inform wetland and floodplain management of the River Murray in South Australia’. Report prepared for the South Australian Murray-Darling Basin Natural Resources Management Board. For further information please contact: Department of Environment and Natural Resources Phone Information Line (08) 8204 1910, or see SA White Pages for your local Department of Environment and Natural Resources office. Online information available at: http://www.environment.sa.gov.au Permissive Licence © State of South Australia through the Department of Environment and Natural Resources. You may copy, distribute, display, download and otherwise freely deal with this publication for any purpose subject to the conditions that you (1) attribute the Department as the copyright owner of this publication and that (2) you obtain the prior written consent of the Department of Environment and Natural Resources if you wish to modify the work or offer the publication for sale or otherwise use it or any part of it for a commercial purpose. Written requests for permission should be addressed to: Design and Production Manager Department of Environment and Natural Resources GPO Box 1047 Adelaide SA 5001 Disclaimer While reasonable efforts have been made to ensure the contents of this publication are factually correct, the Department of Environment and Natural Resources makes no representations and accepts no responsibility for the accuracy, completeness or fitness for any particular purpose of the contents, and shall not be liable for any loss or damage that may be occasioned directly or indirectly through the use of or reliance on the contents of this publication. -
Dwarf Thistle, Cirsi
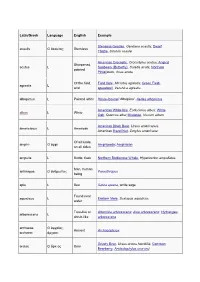
Latin/Greek Language English Example Stemless Gentian, Gentiana acaulis; Dwarf acaulis G ἄκαυλος Stemless Thistle, Cirsium acaule American Crocodile, Crocodylus acutus; Angled Sharpened, acutus L Sunbeam (Butterfly), Curetis acuta; Northern pointed Pintailduck, Anas acuta Of the field, Field Vole, Microtus agrestis; Green Field- agrestis L wild speedwell, Veronica agrestis albopictus L Painted white Hosta fortunei 'Albopicta', Aedes albopictus American White Ibis, Eudocimus albus; White albus L White Oak, Quercus alba; Mistletoe, Viscum album American Black Bear, Ursus americanus; americanus L American American Hazel Nut, Corylus americana Of all kinds, amphi- G ἀμφί Amphipoda; Amphibian on all sides ampulla L Bottle, flask Northern Bottlenose Whale, Hyperoodon ampullatus Man, human anthropos G ἄνθρωπος Paranthropus being apis L Bee Salvia apiana, white sage Found near aquaticus L Eastern Mole, Scalopus aquaticus water Tree-like or Artemisia arborescens; Aloe arborescens; Hydrangea arborescens L shrub-like arborescens archaeos, G ἀρχαῖος, Ancient Archaeopteryx archaeo- ἀρχαιο- Grizzly Bear, Ursus arctos horribilis; Common arctos G ἄρκτος Bear Bearberry, Arctostaphylos uva-ursi argentatus L Silvery Herring Gull, Larus argentatus arthron G ἄρθρον Joint Arthropoda arvensis L In the field Skylark, Alauda arvensis astron, astro-, G ἄστρον, Star Starfish (class), Asteroidea astero- ἀστρο-, ἀστερο- Acer palmatum 'Atropurpureum'; Berberis atropurpureum L Deep purple thunbergii f. atropurpurea Daphne odora 'Aureomarginata'; Taxus aureomarginata -

Common Poultry Diseases 1 G
PS47 Common Poultry Diseases 1 G. D. Butcher, J. P. Jacob, and F. B. Mather2 Respiratory Diseases respiratory distress by obstructing the upper air passages. Chickens may be affected with either or both forms of fowl There are many common and important diseases which can pox at one time. affect the respiratory system (air passages, lungs, air sacs) of poultry (see Table 1). Poultry refers to birds that people Transmission: Fowl pox is transmitted by direct contact keep for their use and generally includes the chicken, between infected and susceptible birds or by mosquitos. turkey, duck, goose, quail, pheasant, pigeon, guinea fowl, Virus-containing scabs also can be sloughed from affected pea fowl, ostrich, emu, and rhea. Due to modern systems birds and serve as a source of infection. The virus can of management, usually with high poultry densities, these enter the blood stream through the eye, skin wounds, or diseases are able to readily spread. respiratory tract. Mosquitos become infected from feeding on birds with fowl pox in their blood stream. There is Fowl Pox some evidence that the mosquito remains infective for life. Synonyms: chicken pox (not to be confused with chicken Mosquitos are the primary reservoir and spreaders of fowl pox in humans; the human disease does not affect poultry pox on poultry ranges. Several species of mosquito can and vice versa), sore head, avian diphtheria, bird pox transmit fowl pox. Often mosquitos winter-over in poultry houses so, outbreaks can occur during winter and early Species affected: Most poultry—chickens, turkeys, pheas- spring. ants, quail, ducks, psittacine, and ratites—of all ages are susceptible. -

Adenoviral Vector-Based Vaccine Platforms for Developing the Next Generation of Influenza Vaccines
Review Adenoviral Vector-Based Vaccine Platforms for Developing the Next Generation of Influenza Vaccines Ekramy E. Sayedahmed 1 , Ahmed Elkashif 1, Marwa Alhashimi 1, Suryaprakash Sambhara 2,* and Suresh K. Mittal 1,* 1 Department of Comparative Pathobiology, Purdue Institute for Immunology, Inflammation and Infectious Disease, Purdue University Center for Cancer Research, College of Veterinary Medicine, Purdue University, West Lafayette, IN 47907, USA; [email protected] (E.E.S.); [email protected] (A.E.); [email protected] (M.A.) 2 Influenza Division, Centers for Disease Control and Prevention, Atlanta, GA 30333, USA * Correspondence: [email protected] (S.S.); [email protected] (S.K.M.) Received: 2 August 2020; Accepted: 17 September 2020; Published: 1 October 2020 Abstract: Ever since the discovery of vaccines, many deadly diseases have been contained worldwide, ultimately culminating in the eradication of smallpox and polio, which represented significant medical achievements in human health. However, this does not account for the threat influenza poses on public health. The currently licensed seasonal influenza vaccines primarily confer excellent strain-specific protection. In addition to the seasonal influenza viruses, the emergence and spread of avian influenza pandemic viruses such as H5N1, H7N9, H7N7, and H9N2 to humans have highlighted the urgent need to adopt a new global preparedness for an influenza pandemic. It is vital to explore new strategies for the development of effective vaccines for pandemic and seasonal influenza viruses. The new vaccine approaches should provide durable and broad protection with the capability of large-scale vaccine production within a short time. The adenoviral (Ad) vector-based vaccine platform offers a robust egg-independent production system for manufacturing large numbers of influenza vaccines inexpensively in a short timeframe. -

Behavioural Adaptation of a Bird from Transient Wetland Specialist to an Urban Resident
Behavioural Adaptation of a Bird from Transient Wetland Specialist to an Urban Resident John Martin1,2*, Kris French1, Richard Major2 1 Institute for Conservation Biology and Environmental Management, School of Biological Sciences, University of Wollongong, Wollongong, New South Wales, Australia, 2 Terrestrial Ecology, Australian Museum, Sydney, New South Wales, Australia Abstract Dramatic population increases of the native white ibis in urban areas have resulted in their classification as a nuisance species. In response to community and industry complaints, land managers have attempted to deter the growing population by destroying ibis nests and eggs over the last twenty years. However, our understanding of ibis ecology is poor and a question of particular importance for management is whether ibis show sufficient site fidelity to justify site-level management of nuisance populations. Ibis in non-urban areas have been observed to be highly transient and capable of moving hundreds of kilometres. In urban areas the population has been observed to vary seasonally, but at some sites ibis are always observed and are thought to be behaving as residents. To measure the level of site fidelity, we colour banded 93 adult ibis at an urban park and conducted 3-day surveys each fortnight over one year, then each quarter over four years. From the quarterly data, the first year resighting rate was 89% for females (n = 59) and 76% for males (n = 34); this decreased to 41% of females and 21% of males in the fourth year. Ibis are known to be highly mobile, and 70% of females and 77% of males were observed at additional sites within the surrounding region (up to 50 km distant). -

Namibia & the Okavango
Pel’s Fishing Owl - a pair was found on a wooded island south of Shakawe (Jan-Ake Alvarsson) NAMIBIA & THE OKAVANGO 21 SEPTEMBER – 8 OCTOBER 2017 LEADER: STEVE BRAINE For most of the country the previous three years drought had been broken and although too early for the mi- grants we did however do very well with birding generally. We searched and found all the near endemics as well as the endemic Dune Lark. Besides these we also had a new write-in for the trip! In the floodplains after observing a wonderful Pel’s Fishing Owl we travelled down a side channel of the Okavango River to look for Pygmy Geese, we were lucky and came across several pairs before reaching a dried-out floodplain. Four birds flew out of the reedbeds and looked rather different to the normal weavers of which there were many, a closer look at the two remaining birds revealed a beautiful pair of Cuckoo Finches. These we all enjoyed for a brief period before they followed the other birds which had now disappeared into the reedbeds. Very strong winds on three of the birding days made birding a huge challenge to say the least after not finding the rare and difficult Herero Chat we had to make alternate arrangements at another locality later in the trip. The entire tour from the Hosea Kutako International Airport outside the capital Windhoek and returning there nineteen days later delivered 375 species. Out of these, four birds were seen only by the leader, a further three species were heard but not seen. -

Ectoparasites of the Laughing Dove Streptopelia Senegalensis (Linnaeus, 1766) (Aves: Columbidae) in Zaria, Nigeria
Lundiana 9(1):67-71, 2008 © 2009 Instituto de Ciências Biológicas - UFMG ISSN 1676-6180 Ectoparasites of the Laughing Dove Streptopelia senegalensis (Linnaeus, 1766) (Aves: Columbidae) in Zaria, Nigeria 1Lucas K. Adang, 2Sonnie J. Oniye, 2Augustine U. Ezealor, 3Paul A. Abdu, 4Joseph O. Ajanusi & 1Kennedy P. Yoriyo 1 Department of Biological Sciences, Gombe State University, Gombe, Nigeria. E-mail: [email protected] 2 Department of Biological Sciences, 3 Department of Surgery and Medicine, 4 Department of Veterinary Parasitology and Entomology, Ahmadu Bello University, Zaria, Nigeria. Abstract A survey of ectoparasites of the Laughing Dove (Streptopelia senegalensis Linnaeus, 1766) was carried out in Zaria, Nigeria, to determine the prevalence, intensity and mean intensity of infestation. A total of 382 (231 males and 151 females) doves trapped from different locations in Zaria, Nigeria, were examined through plumage brushing. Eighty-eight (23.0%) of the birds were infested by the following six species of ectoparasites: lice – 32 (8.4%) Menopon gallinae Linnaeus, 1758, 37 (9.7%) Columbicola columbae Linnaeus, 1758, and 18(4.7%) Goniodes sp.; flies – 19 (5.0%) Pseudolynchia canariensis Macquart, 1840; ticks – 12 (3.1%) Argas persicus Oken, 1818; and mite: 1 (0.23%) Dermanyssus gallinae (Degeer, 1778). The frequency of single infestations (59 – 15.4%), was higher than that of double (27 – 7.1%) and triple (2 – 0.52%) infestations, though the difference was not statistically significant (p > 0.05). The males had a higher prevalence (55 – 23.8%) than the females (33 – 21.9%). However, this difference was also not significant (p > 0.05). Ectoparasites were collected from the birds through out the year, with highest prevalence (60.0%) in November. -

Avifauna from the Teouma Lapita Site, Efate Island, Vanuatu, Including a New Genus and Species of Megapode
Archived at the Flinders Academic Commons: http://dspace.flinders.edu.au/dspace/ ‘This is the peer reviewed version of the following article: Worthy, T., Hawkins, S., Bedford, S. and Spriggs, M. (2015). Avifauna from the Teouma Lapita Site, Efate Island, Vanuatu, including a new genus and species of megapode. Pacific Science, 69(2) pp. 205-254. which has been published in final form at DOI: http://dx.doi.org/10.2984/69.2.6 Article: http://www.bioone.org/doi/full/10.2984/69.2.6 Journal: http://www.uhpress.hawaii.edu/t-pacific-science Copyright 2015, University of Hawaii Press. Published version of the article is reproduced here with permission from the publisher." Avifauna from the Teouma Lapita Site, Efate Island, Vanuatu, Including a New Genus and Species of Megapode1 Trevor H. Worthy,2,5 Stuart Hawkins,3 Stuart Bedford,4 and Matthew Spriggs 4 Abstract: The avifauna of the Teouma archaeological site on Efate in Vanuatu is described. It derives from the Lapita levels (3,000 – 2,800 ybp) and immedi- ately overlying middens extending to ~2,500 ybp. A total of 30 bird species is represented in the 1,714 identified specimens. Twelve species are new records for the island, which, added to previous records, indicates that minimally 39 land birds exclusive of passerines were in the original avifauna. Three-fourths of the 12 newly recorded species appear to have become extinct by the end of Lapita times, 2,800 ybp. The avifauna is dominated by eight species of columbids (47.5% Minimum Number Individuals [MNI ]) including a large extinct tooth- billed pigeon, Didunculus placopedetes from Tonga, and a giant Ducula sp. -
POULTRY INDUSTRY OVERVIEW Chicken Is a Multi-Billion Dollar Business (For a Few)
POULTRY INDUSTRY OVERVIEW Chicken is a multi-billion dollar business (for a few) America’s favorite meat is a gold mine. In 2014, chicken generated revenues of $ 32.7 billion in the US alone. The three biggest companies control almost 60% of the production, concentrating power in the hands of a few corporations. Poultry companies are called Top Five Poultry Companies INTEGRATORS and they CONTRACT Soy Bean (Ready-To-Cook chicken - mil lb/week) farmers to raise their chickens Tyson Foods 176.64 Pilgrim’s Pride INTEGRATED =owned by the company Corn/Soybean 138.36 The integrator purchases the grains Perdue 56.49 to produce feed Sanderson Farms 58.80 Koch Foods 48.00 Others 159.79 Breeder Company Feed Mill Breeds are developed and owned by 27.6% The integrator creates their own secret feed mixes a firm, oen a subsidiary that they deliver to the farmers 21.7% SUBSIDIARY INTEGRATED 25% 9% 9.2% 7.5% Hatchery Chicks are hatched and sent to broiler farmers Breeder Farm These farms raise chickens who lay INTEGRATED eggs for the hatchery UNDER CONTRACT Broiler Farm Slaughter House Chicks and feed that are owned by the company are delivered to broiler farmers In 2014, chicken farmers raised UNDER CONTRACT Tyson Foods 8.5 billion chickens in the USA 33Processing Plants 33Million/week There are almost 30,000 poultry farmers in the US Pilgrim’s Pride 97% of the chicken we eat is produced under contract 24 Processing Plants 28Million/week INTEGRATED UNDER CONTRACT Tyson Foods produced 1.8 billion chickens Further Processing Pilgrim’s Pride produced 1.5 billion chickens Raw carcasses are turned into frozen or pre-cooked products and packaged for consumers Chicks are raised into slaughter INTEGRATED weight broiler chickens in less than 5 weeks MON TUE WED THU FRI SAT SUN Own Brand Food Services Pilgrim’s Pride as well as most of the poultry companies have contracts with retailers and food services to provide a predictable amount of products with a Many companies like Tyson own their own brands that standardized quality.