Cerebellar Granule Cell-Specific and Inducible Expression of Cre Recombinase in the Mouse
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

System Inflammation Macrophages/Microglia in Central Nervous Brain Dendritic Cells
Brain Dendritic Cells and Macrophages/Microglia in Central Nervous System Inflammation This information is current as Hans-Georg Fischer and Gaby Reichmann of September 25, 2021. J Immunol 2001; 166:2717-2726; ; doi: 10.4049/jimmunol.166.4.2717 http://www.jimmunol.org/content/166/4/2717 Downloaded from References This article cites 51 articles, 28 of which you can access for free at: http://www.jimmunol.org/content/166/4/2717.full#ref-list-1 Why The JI? Submit online. http://www.jimmunol.org/ • Rapid Reviews! 30 days* from submission to initial decision • No Triage! Every submission reviewed by practicing scientists • Fast Publication! 4 weeks from acceptance to publication *average by guest on September 25, 2021 Subscription Information about subscribing to The Journal of Immunology is online at: http://jimmunol.org/subscription Permissions Submit copyright permission requests at: http://www.aai.org/About/Publications/JI/copyright.html Email Alerts Receive free email-alerts when new articles cite this article. Sign up at: http://jimmunol.org/alerts The Journal of Immunology is published twice each month by The American Association of Immunologists, Inc., 1451 Rockville Pike, Suite 650, Rockville, MD 20852 Copyright © 2001 by The American Association of Immunologists All rights reserved. Print ISSN: 0022-1767 Online ISSN: 1550-6606. Brain Dendritic Cells and Macrophages/Microglia in Central Nervous System Inflammation1 Hans-Georg Fischer2 and Gaby Reichmann Microglia subpopulations were studied in mouse experimental autoimmune encephalomyelitis and toxoplasmic encephalitis. CNS .inflammation was associated with the proliferation of CD11b؉ brain cells that exhibited the dendritic cell (DC) marker CD11c These cells constituted up to 30% of the total CD11b؉ brain cell population. -

Mixed Electrical–Chemical Synapses in Adult Rat Hippocampus Are Primarily Glutamatergic and Coupled by Connexin-36
ORIGINAL RESEARCH ARTICLE published: 15 May 2012 NEUROANATOMY doi: 10.3389/fnana.2012.00013 Mixed electrical–chemical synapses in adult rat hippocampus are primarily glutamatergic and coupled by connexin-36 Farid Hamzei-Sichani 1,2,3‡, Kimberly G. V. Davidson4‡,ThomasYasumura4,William G. M. Janssen3, Susan L.Wearne 4#, Patrick R. Hof 3, Roger D.Traub 5†, Rafael Gutiérrez 6, Ole P.Ottersen7 and John E. Rash4,8* 1 Department of Neurosurgery, Mount Sinai School of Medicine, New York, NY, USA 2 Program in Neural and Behavioral Science, Downstate Medical Center, State University of New York, Brooklyn, NY, USA 3 Fishberg Department of Neuroscience, Friedman Brain Institute, Mount Sinai School of Medicine, New York, NY, USA 4 Department of Biomedical Sciences, Colorado State University, Fort Collins, CO, USA 5 Department of Physiology and Pharmacology, Downstate Medical Center, State University of New York, Brooklyn, NY, USA 6 Department of Pharmacobiology, Centro de Investigación y Estudios Avanzados del Instituto Politécnico Nacional, México D.F. 7 Centre for Molecular Biology and Neuroscience, University of Oslo, Oslo, Norway 8 Program in Molecular, Cellular, and Integrative Neurosciences, Colorado State University, Fort Collins, CO, USA Edited by: Dendrodendritic electrical signaling via gap junctions is now an accepted feature of neu- Ryuichi Shigemoto, National Institute ronal communication in mammalian brain, whereas axodendritic and axosomatic gap junc- for Physiological Sciences, Japan tions have rarely been described. We present ultrastructural, immunocytochemical, and Reviewed by: Javier DeFelipe, Cajal Institute, Spain dye-coupling evidence for “mixed” (electrical/chemical) synapses on both principal cells Richard J. Weinberg, University of and interneurons in adult rat hippocampus. -

Deconstructing Spinal Interneurons, One Cell Type at a Time Mariano Ignacio Gabitto
Deconstructing spinal interneurons, one cell type at a time Mariano Ignacio Gabitto Submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy under the Executive Committee of the Graduate School of Arts and Sciences COLUMBIA UNIVERSITY 2016 © 2016 Mariano Ignacio Gabitto All rights reserved ABSTRACT Deconstructing spinal interneurons, one cell type at a time Mariano Ignacio Gabitto Abstract Documenting the extent of cellular diversity is a critical step in defining the functional organization of the nervous system. In this context, we sought to develop statistical methods capable of revealing underlying cellular diversity given incomplete data sampling - a common problem in biological systems, where complete descriptions of cellular characteristics are rarely available. We devised a sparse Bayesian framework that infers cell type diversity from partial or incomplete transcription factor expression data. This framework appropriately handles estimation uncertainty, can incorporate multiple cellular characteristics, and can be used to optimize experimental design. We applied this framework to characterize a cardinal inhibitory population in the spinal cord. Animals generate movement by engaging spinal circuits that direct precise sequences of muscle contraction, but the identity and organizational logic of local interneurons that lie at the core of these circuits remain unresolved. By using our Sparse Bayesian approach, we showed that V1 interneurons, a major inhibitory population that controls motor output, fractionate into diverse subsets on the basis of the expression of nineteen transcription factors. Transcriptionally defined subsets exhibit highly structured spatial distributions with mediolateral and dorsoventral positional biases. These distinctions in settling position are largely predictive of patterns of input from sensory and motor neurons, arguing that settling position is a determinant of inhibitory microcircuit organization. -

Canine Dorsal Root Ganglia Satellite Glial Cells Represent an Exceptional Cell Population with Astrocytic and Oligodendrocytic P
www.nature.com/scientificreports OPEN Canine dorsal root ganglia satellite glial cells represent an exceptional cell population with astrocytic and Received: 17 August 2017 Accepted: 6 October 2017 oligodendrocytic properties Published: xx xx xxxx W. Tongtako1,2, A. Lehmbecker1, Y. Wang1,2, K. Hahn1,2, W. Baumgärtner1,2 & I. Gerhauser 1 Dogs can be used as a translational animal model to close the gap between basic discoveries in rodents and clinical trials in humans. The present study compared the species-specifc properties of satellite glial cells (SGCs) of canine and murine dorsal root ganglia (DRG) in situ and in vitro using light microscopy, electron microscopy, and immunostainings. The in situ expression of CNPase, GFAP, and glutamine synthetase (GS) has also been investigated in simian SGCs. In situ, most canine SGCs (>80%) expressed the neural progenitor cell markers nestin and Sox2. CNPase and GFAP were found in most canine and simian but not murine SGCs. GS was detected in 94% of simian and 71% of murine SGCs, whereas only 44% of canine SGCs expressed GS. In vitro, most canine (>84%) and murine (>96%) SGCs expressed CNPase, whereas GFAP expression was diferentially afected by culture conditions and varied between 10% and 40%. However, GFAP expression was induced by bone morphogenetic protein 4 in SGCs of both species. Interestingly, canine SGCs also stimulated neurite formation of DRG neurons. These fndings indicate that SGCs represent an exceptional, intermediate glial cell population with phenotypical characteristics of oligodendrocytes and astrocytes and might possess intrinsic regenerative capabilities in vivo. Since the discovery of glial cells over a century ago, substantial progress has been made in understanding the origin, development, and function of the diferent types of glial cells in the central nervous system (CNS) and peripheral nervous system (PNS)1. -

Control of Cerebellar Granule Cell Output by Sensory-Evoked Golgi Cell Inhibition
Control of cerebellar granule cell output by sensory-evoked Golgi cell inhibition Ian Duguid1,2,3, Tiago Branco1,4, Paul Chadderton5, Charlotte Arlt, Kate Powell6, and Michael Häusser3 Wolfson Institute for Biomedical Research and Department of Neuroscience, Physiology, and Pharmacology, University College London, London WC1E 6BT, United Kingdom Edited by Masao Ito, RIKEN Brain Science Institute, Wako, Japan, and approved September 1, 2015 (received for review May 25, 2015) Classical feed-forward inhibition involves an excitation–inhibition Results sequence that enhances the temporal precision of neuronal re- Sensory-Evoked Phasic and Spillover Golgi Cell Inhibition Precedes sponses by narrowing the window for synaptic integration. In Mossy Fiber Excitation in Granule Cells. Cerebellar granule cells the input layer of the cerebellum, feed-forward inhibition is thought receive direct phasic and indirect or “spillover” GABAergic in- to preserve the temporal fidelity of granule cell spikes during mossy put from Golgi cells (6, 16, 20, 21). To investigate the temporal fiber stimulation. Although this classical feed-forward inhibitory cir- dynamics of sensory-evoked inhibition in vivo, we recorded cuit has been demonstrated in vitro, the extent to which inhibition spontaneous and sensory-evoked excitatory (Vhold = −70 mV) shapes granule cell sensory responses in vivo remains unresolved. and inhibitory (Vhold = 0 mV) currents from the same granule Here we combined whole-cell patch-clamp recordings in vivo and cells in Crus II (Fig. 1 A–D). Granule cells were identified based dynamic clamp recordings in vitro to directly assess the impact of on their characteristic electrophysiological properties (Table S1), Golgi cell inhibition on sensory information transmission in the depth from the pial surface (>250 μm), and morphology (Fig. -

Comparative Morphology of Gigantopyramidal Neurons in Primary Motor Cortex Across Mammals
Received: 23 August 2017 | Revised: 19 October 2017 | Accepted: 24 October 2017 DOI: 10.1002/cne.24349 The Journal of RESEARCH ARTICLE Comparative Neurology Comparative morphology of gigantopyramidal neurons in primary motor cortex across mammals Bob Jacobs1 | Madeleine E. Garcia1 | Noah B. Shea-Shumsky1 | Mackenzie E. Tennison1 | Matthew Schall1 | Mark S. Saviano1 | Tia A. Tummino1 | Anthony J. Bull2 | Lori L. Driscoll1 | Mary Ann Raghanti3 | Albert H. Lewandowski4 | Bridget Wicinski5 | Hong Ki Chui1 | Mads F. Bertelsen6 | Timothy Walsh7 | Adhil Bhagwandin8 | Muhammad A. Spocter8,9,10 | Patrick R. Hof5 | Chet C. Sherwood11 | Paul R. Manger8 1Laboratory of Quantitative Neuromorphology, Neuroscience Program, Colorado College, Colorado Springs, Colorado 2Human Biology and Kinesiology, Colorado College, Colorado Springs, Colorado 3Department of Anthropology and School of Biomedical Sciences, Kent State University, Kent, Ohio 4Cleveland Metroparks Zoo, Cleveland, Ohio 5Fishberg Department of Neuroscience and Friedman Brain Institute, Icahn School of Medicine at Mount Sinai, New York, New York 6Center for Zoo and Wild Animal Health, Copenhagen Zoo, Fredericksberg, Denmark 7Smithsonian National Zoological Park, Washington, District of Columbia 8School of Anatomical Sciences, Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa 9Department of Anatomy, Des Moines University, Des Moines, Iowa 10Biomedical Sciences, College of Veterinary Medicine, Iowa State University, Ames, Iowa 11Department of Anthropology and Center for the Advanced Study of Human Paleobiology, The George Washington University, Washington, District of Columbia Correspondence Bob Jacobs, Ph.D., Laboratory of Abstract Quantitative Neuromorphology, Gigantopyramidal neurons, referred to as Betz cells in primates, are characterized by large somata Neuroscience Program, Colorado College, and extensive basilar dendrites. Although there have been morphological descriptions and draw- 14 E. -

Astrocytes: a Driving Force in Brain Signaling and Encephalopathies Aleksandar Jeremic Iowa State University
Iowa State University Capstones, Theses and Retrospective Theses and Dissertations Dissertations 2001 Astrocytes: a driving force in brain signaling and encephalopathies Aleksandar Jeremic Iowa State University Follow this and additional works at: https://lib.dr.iastate.edu/rtd Part of the Neuroscience and Neurobiology Commons, and the Neurosciences Commons Recommended Citation Jeremic, Aleksandar, "Astrocytes: a driving force in brain signaling and encephalopathies " (2001). Retrospective Theses and Dissertations. 648. https://lib.dr.iastate.edu/rtd/648 This Dissertation is brought to you for free and open access by the Iowa State University Capstones, Theses and Dissertations at Iowa State University Digital Repository. It has been accepted for inclusion in Retrospective Theses and Dissertations by an authorized administrator of Iowa State University Digital Repository. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand comer and continuing from left to right in equal sections with small overlaps. -

A Single-Neuron: Current Trends and Future Prospects
cells Review A Single-Neuron: Current Trends and Future Prospects Pallavi Gupta 1, Nandhini Balasubramaniam 1, Hwan-You Chang 2, Fan-Gang Tseng 3 and Tuhin Subhra Santra 1,* 1 Department of Engineering Design, Indian Institute of Technology Madras, Tamil Nadu 600036, India; [email protected] (P.G.); [email protected] (N.B.) 2 Department of Medical Science, National Tsing Hua University, Hsinchu 30013, Taiwan; [email protected] 3 Department of Engineering and System Science, National Tsing Hua University, Hsinchu 30013, Taiwan; [email protected] * Correspondence: [email protected] or [email protected]; Tel.: +91-044-2257-4747 Received: 29 April 2020; Accepted: 19 June 2020; Published: 23 June 2020 Abstract: The brain is an intricate network with complex organizational principles facilitating a concerted communication between single-neurons, distinct neuron populations, and remote brain areas. The communication, technically referred to as connectivity, between single-neurons, is the center of many investigations aimed at elucidating pathophysiology, anatomical differences, and structural and functional features. In comparison with bulk analysis, single-neuron analysis can provide precise information about neurons or even sub-neuron level electrophysiology, anatomical differences, pathophysiology, structural and functional features, in addition to their communications with other neurons, and can promote essential information to understand the brain and its activity. This review highlights various single-neuron models and their behaviors, followed by different analysis methods. Again, to elucidate cellular dynamics in terms of electrophysiology at the single-neuron level, we emphasize in detail the role of single-neuron mapping and electrophysiological recording. We also elaborate on the recent development of single-neuron isolation, manipulation, and therapeutic progress using advanced micro/nanofluidic devices, as well as microinjection, electroporation, microelectrode array, optical transfection, optogenetic techniques. -
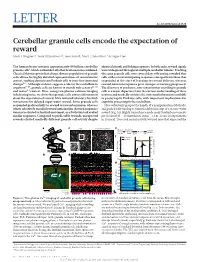
Cerebellar Granule Cells Encode the Expectation of Reward Mark J
LETTER doi:10.1038/nature21726 Cerebellar granule cells encode the expectation of reward Mark J. Wagner1*, Tony Hyun Kim1,2*, Joan Savall1, Mark J. Schnitzer1,3 & Liqun Luo1 The human brain contains approximately 60 billion cerebellar identical stimuli and licking responses. In both tasks, reward signals granule cells1, which outnumber all other brain neurons combined. were widespread throughout multiple cerebellar lobules. Tracking Classical theories posit that a large, diverse population of granule the same granule cells over several days of learning revealed that cells allows for highly detailed representations of sensorimotor cells with reward-anticipating responses emerged from those that context, enabling downstream Purkinje cells to sense fine contextual responded at the start of learning to reward delivery, whereas changes2–6. Although evidence suggests a role for the cerebellum in reward-omission responses grew stronger as learning progressed. cognition7–10, granule cells are known to encode only sensory11–13 The discovery of predictive, non-sensorimotor encoding in granule and motor14 context. Here, using two-photon calcium imaging cells is a major departure from the current understanding of these in behaving mice, we show that granule cells convey information neurons and markedly enriches the contextual information available about the expectation of reward. Mice initiated voluntary forelimb to postsynaptic Purkinje cells, with important implications for movements for delayed sugar-water reward. Some granule cells cognitive processing in the cerebellum. responded preferentially to reward or reward omission, whereas Mice voluntarily grasped the handle of a manipulandum (Methods) others selectively encoded reward anticipation. Reward responses and pushed it forward up to 8 mm for delayed receipt of a sucrose-water were not restricted to forelimb movement, as a Pavlovian task evoked reward (Fig. -

Selective Gene Expression in Brain Microglia Mediated Via Adeno-Associated Virus Type 2 and Type 5 Vectors
Gene Therapy (2003) 10, 657–667 & 2003 Nature Publishing Group All rights reserved 0969-7128/03 $25.00 www.nature.com/gt RESEARCH ARTICLE Selective gene expression in brain microglia mediated via adeno-associated virus type 2 and type 5 vectors M Cucchiarini1,w, XL Ren1, G Perides2 and EF Terwilliger1 1Division of Experimental Medicine, Harvard Institutes of Medicine and Beth Israel Deaconess Medical Center, Boston, MA, USA; and 2Department of Surgery, Beth Israel Deaconess Medical Center, and Harvard Medical School, Boston, MA, USA Microglia represent a crucial cell population in the central examined, an element derived from the gene for the murine nervous system, participating in the regulation and surveil- macrophage marker F4/80 was the most discriminating for lance of physiological processes as well as playing key roles microglia. Gene expression from vectors controlled by this in the etiologies of several major brain disorders. The ability element was highly selective for microglia, both in vitro and in to target gene transfer vehicles selectively to microglia would vivo. To our knowledge, this is the first demonstration of provide a powerful new approach to investigations of selective expression of transferred genes in microglia using mechanisms regulating brain pathologies, as well as enable AAV-derived vectors, as well as the first utilization of the development of novel therapeutic strategies. In this recombinant AAV-5 vectors in any macrophage lineage. study, we evaluate the feasibility of specifically and efficiently These results provide strong encouragement for the applica- targeting microglia relative to other brain cells, using vectors tion of these vectors and this approach for delivering based on two different serotypes of adeno-associated virus therapeutic and other genes selectively to microglia. -

Microglia Lifelong Patrolling Immune Cells of the Brain
Progress in Neurobiology 179 (2019) 101614 Contents lists available at ScienceDirect Progress in Neurobiology journal homepage: www.elsevier.com/locate/pneurobio Review article Microglia: Lifelong patrolling immune cells of the brain T ⁎ Ukpong B. Eyoa,b,c, Long-Jun Wua,d,e, a Department of Neurology, Mayo Clinic, Rochester, MN 55905, USA b Center for Brain Immunology and Glia (BIG), University of Virginia, Charlottesville, VA, USA c Department of Neuroscience, University of Virginia, Charlottesville, VA, USA d Department of Neuroscience, Mayo Clinic, Jacksonville, FL 32224, USA e Department of Immunology, Mayo Clinic, Rochester, MN 55905, USA ARTICLE INFO ABSTRACT Keywords: Microglial cells are the predominant parenchymal immune cell of the brain. Recent evidence suggests that like Microglia peripheral immune cells, microglia patrol the brain in health and disease. Reviewing these data, we first examine Surveillance the evidence that microglia invade the brain mesenchyme early in embryonic development, establish residence Microglial landscape therein, proliferate and subsequently maintain their numbers throughout life. We, then, summarize established Neuroimmune interaction and novel evidence for microglial process surveillance in the healthy and injured brain. Finally, we discuss Epilepsy emerging evidence for microglial cell body dynamics that challenge existing assumptions of their sessile nature. We conclude that microglia are long-lived immune cells that patrol the brain through both cell body and process movements. This recognition -

Drosophila Glia: Models for Human Neurodevelopmental and Neurodegenerative Disorders
International Journal of Molecular Sciences Review Drosophila Glia: Models for Human Neurodevelopmental and Neurodegenerative Disorders Taejoon Kim, Bokyeong Song and Im-Soon Lee * Department of Biological Sciences, Center for CHANS, Konkuk University, Seoul 05029, Korea; [email protected] (T.K.); [email protected] (B.S.) * Correspondence: [email protected] Received: 31 May 2020; Accepted: 7 July 2020; Published: 9 July 2020 Abstract: Glial cells are key players in the proper formation and maintenance of the nervous system, thus contributing to neuronal health and disease in humans. However, little is known about the molecular pathways that govern glia–neuron communications in the diseased brain. Drosophila provides a useful in vivo model to explore the conserved molecular details of glial cell biology and their contributions to brain function and disease susceptibility. Herein, we review recent studies that explore glial functions in normal neuronal development, along with Drosophila models that seek to identify the pathological implications of glial defects in the context of various central nervous system disorders. Keywords: glia; glial defects; Drosophila models; CNS disorders 1. Introduction Glial cells perform many important functions that are essential for the proper development and maintenance of the nervous system [1]. During development, glia maintain neuronal cell numbers and engulf unnecessary cells and projections, correctly shaping neural circuits. In comparison, glial cells in the adult brain provide metabolic sustenance and critical immune support. Thus, the dysfunction of glial activity contributes to various central nervous system (CNS) disorders in humans at different stages of life [2]. Accordingly, the need for research regarding the initiation as well as the progression of disorders associated with glial cell dysfunction is increasing.