Us and Life History of the Three Assessed Mussels
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Native Freshwater Mussels
Native Freshwater Mussels Freshwater mussels, sometimes called clams, have always been and continue to be, an important food source for muskrat, minks, FACT SHEET raccoons, otters, fishes, and some birds such as herons. Historically, written by John Tautin Native Americans not only ate mussels but also used the shells for utensils, tools, and to make jewelry. Between the late 1800s and mid-1900s, shells were harvested to supply a multi- fluted shell million dollar pearl button industry. However, with the Lasmigona costata invention and widespread use of plastics during the 1940-50s, the pearl button industry collapsed. But by the 1950s the Japanese found a new use for mussel shells in cultured pearl production. e shells are cut and finished into beads and inserted into oysters to serve as nuclei for pearls. Still today, thousands of tons of mussel shells (especially Washboard, Mapleleaf, and ree-ridge mussels) are exported from the United States to Japan for this purpose. Elktoe Worldwide, there are about one thousand species of mussels. Mussels Alasmidonta marginata can be found on every continent but Antarctica. While the entire continent of Europe only has eight different species of mussels, there are twenty-five different species of mussels in French Creek, and about three-hundred in the United States. ese important animals are threatened, however. Today, over half of the species of mussel in the Midwest are threatened or endangered. In the French Creek watershed, thirteen species of mussels are listed as endan- gered or threatened in Pennsylvania. Four species (the northern riffleshell, kidney shell and clubshell, rayed bean, and snuffbox) are endangered at the federal level. -

Tennessee's Extinct Species
Tennessee's Extinct Species The following species Birds: once occurred in Carolina parakeet Conuropsis carolinensis Ectopistes migratorius Tennessee and are now Passenger pigeon believed to be extinct. Mammals: Following this list are two Eastern elk species descriptions-one Fishes: describing the Carolina Harelip sucker parakeet and another describing the extinct Mussels: Acornshell Epioblasma haysiana freshwater mussels Angled riffleshell Epioblasma biemarginata of Tennessee. Cumberland leafshell Epioblasma stewardsoni Leafshell Epioblasma flexuosa Narrowcat's paw Epioblasma lenoir Rough rockshell Quadrula tuberosa Round combshell Epioblasma personata Sugarspoon Epioblasma arcaeformis Tennessee riffleshell Epioblasma propinqua Carolina Parakeet Status Habitat The Carolina parakeet is an The Carolina parakeet was found Learn rrwreabout extinct species. in riverine forests, cypress swamps, Tennessee's diverse and other woodlands over much of Description the Eastern and Midwest Regions of ecosyster.n3.Su~ort The Carolina parakeet was a the United States. It was the only conservation in your small parrot, about 12inches in parrot native to the United States. community and state! length. Its head was lemon yellow, The parakeets rested at night in with an orange forehead and cheeks. groups, with as many as 30 birds The rest of its body was green. Its sleeping inside one hollowtree, while legs and beak were pale pinkish- others would hang on the outside. white. These curious birds lived and Nests were placed in hollowtrees, traveled in flocks. and three to five white eggs were laid. Up to 50 nests were often crowded into one tree. Role in the Ecosystem Carolina parakeets enjoyed a variety of different foods-apples, peaches, mulberries, pecans, grapes, dogwood fruit, and grains. -

ECOLOGY of NORTH AMERICAN FRESHWATER FISHES
ECOLOGY of NORTH AMERICAN FRESHWATER FISHES Tables STEPHEN T. ROSS University of California Press Berkeley Los Angeles London © 2013 by The Regents of the University of California ISBN 978-0-520-24945-5 uucp-ross-book-color.indbcp-ross-book-color.indb 1 44/5/13/5/13 88:34:34 AAMM uucp-ross-book-color.indbcp-ross-book-color.indb 2 44/5/13/5/13 88:34:34 AAMM TABLE 1.1 Families Composing 95% of North American Freshwater Fish Species Ranked by the Number of Native Species Number Cumulative Family of species percent Cyprinidae 297 28 Percidae 186 45 Catostomidae 71 51 Poeciliidae 69 58 Ictaluridae 46 62 Goodeidae 45 66 Atherinopsidae 39 70 Salmonidae 38 74 Cyprinodontidae 35 77 Fundulidae 34 80 Centrarchidae 31 83 Cottidae 30 86 Petromyzontidae 21 88 Cichlidae 16 89 Clupeidae 10 90 Eleotridae 10 91 Acipenseridae 8 92 Osmeridae 6 92 Elassomatidae 6 93 Gobiidae 6 93 Amblyopsidae 6 94 Pimelodidae 6 94 Gasterosteidae 5 95 source: Compiled primarily from Mayden (1992), Nelson et al. (2004), and Miller and Norris (2005). uucp-ross-book-color.indbcp-ross-book-color.indb 3 44/5/13/5/13 88:34:34 AAMM TABLE 3.1 Biogeographic Relationships of Species from a Sample of Fishes from the Ouachita River, Arkansas, at the Confl uence with the Little Missouri River (Ross, pers. observ.) Origin/ Pre- Pleistocene Taxa distribution Source Highland Stoneroller, Campostoma spadiceum 2 Mayden 1987a; Blum et al. 2008; Cashner et al. 2010 Blacktail Shiner, Cyprinella venusta 3 Mayden 1987a Steelcolor Shiner, Cyprinella whipplei 1 Mayden 1987a Redfi n Shiner, Lythrurus umbratilis 4 Mayden 1987a Bigeye Shiner, Notropis boops 1 Wiley and Mayden 1985; Mayden 1987a Bullhead Minnow, Pimephales vigilax 4 Mayden 1987a Mountain Madtom, Noturus eleutherus 2a Mayden 1985, 1987a Creole Darter, Etheostoma collettei 2a Mayden 1985 Orangebelly Darter, Etheostoma radiosum 2a Page 1983; Mayden 1985, 1987a Speckled Darter, Etheostoma stigmaeum 3 Page 1983; Simon 1997 Redspot Darter, Etheostoma artesiae 3 Mayden 1985; Piller et al. -
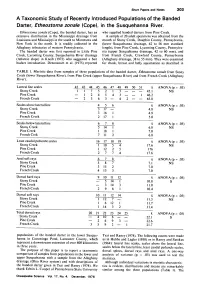
A Taxonomic Study of Recently Introduced Populations of The
Short Papers and Notes 303 A TaxonomicStudy of RecentlyIntroduced Populations of the Banded Darter,Etheostoma zonale (Cope), in the Susquehanna River. Etheostoma zonale (Cope), the banded darter, has an who supplied banded darters from Pine Creek. extensive distribution in the Mississippi drainage from A sample of 20 adult specimens was obtained from the Louisiana and Mississippi in the south to Minnesota and mouth of Stony Creek, Dauphin County, Pennsylvania, New York in the north. It is readily collected in the (lower Susquehanna drainage, 42 to 56 mm standard Allegheny tributaries of western Pennsylvania. length), from Pine Creek, Lycoming County, Pennsylva- The banded darter was first reported in Little Pine nia (upper Susquehanna drainage, 42 to 60 mm), and Creek, Lycoming County, Susquehanna River drainage from French Creek, Crawford County, Pennsylvania (Atlantic slope) in Kneib (1972) who suggested a bait (Allegheny drainage, 38 to 55 mm). They were examined bucket introduction. Denoncourt et al. (1975) reported for cheek, breast and belly squamation as described in TABLE 1. Meristic data from samples of three populations of the banded darter, Etheostoma zonale from Stony Creek (lower Susquehanna River), from Pine Creek (upper Susquehanna River) and from French Creek (Allegheny River). Lateral line scales 42 43 44 45 46 47 48 49 50 51 k ANOVA (p = .05) Stony Creek 1 1 7 5 2 1 3 - -- 45.1 NS Pine Creek - 1 2 4 5 4 3 - - 1 46.2 French Creek - 2 3 6 3 - 4 2 - - 45.8 Scales above lateral line 4 5 6 R ANOVA (p = .05) Stony Creek 3 -

Restoring the Endangered Oyster Mussel (Epioblasma Capsaeformis) to the Upper Clinch River, Virginia: an Evaluation of Population Restoration Techniques Caitlin S
RESEARCH ARTICLE Restoring the endangered oyster mussel (Epioblasma capsaeformis) to the upper Clinch River, Virginia: an evaluation of population restoration techniques Caitlin S. Carey1,2,3,JessW.Jones4, Robert S. Butler5, Eric M. Hallerman6 From 2005 to 2011, the federally endangered freshwater mussel Epioblasma capsaeformis (oyster mussel) was reintroduced at three sites in the upper Clinch River, Virginia, using four release techniques. These release techniques were (1) translocation of adults (site 1, n = 1418), (2) release of laboratory-propagated sub-adults (site 1, n = 2851), (3) release of 8-week-old laboratory-propagated juveniles (site 2, n = 9501), and (4) release of artificially infested host fishes (site 3, n = 1116 host fishes). These restoration efforts provided a unique research opportunity to compare the effectiveness of techniques used to reestablish populations of extirpated and declining species. We evaluated the relative success of these four population restoration approaches via monitoring at each release site (2011–2012) using systematic 0.25-m2 quadrat sampling to estimate abundance and post-release survival. Abundances of translocated adult and laboratory-propagated sub-adult E. capsaeformis at site 1 ranged 577–645 and 1678–1700 individuals, respectively, signifying successful settlement and high post-release survival. Two untagged individuals (29.1 and 27.3 mm) were observed, indicating that recruitment is occurring at site 1. No E. capsaeformis were found at sites where 8-week-old laboratory-propagated juveniles (site 2) and artificially infested host fishes (site 3) were released. Our results indicate that translocations of adults and releases of laboratory-propagated sub-adults were the most effective population restoration techniques for E. -

Louisiana's Animal Species of Greatest Conservation Need (SGCN)
Louisiana's Animal Species of Greatest Conservation Need (SGCN) ‐ Rare, Threatened, and Endangered Animals ‐ 2020 MOLLUSKS Common Name Scientific Name G‐Rank S‐Rank Federal Status State Status Mucket Actinonaias ligamentina G5 S1 Rayed Creekshell Anodontoides radiatus G3 S2 Western Fanshell Cyprogenia aberti G2G3Q SH Butterfly Ellipsaria lineolata G4G5 S1 Elephant‐ear Elliptio crassidens G5 S3 Spike Elliptio dilatata G5 S2S3 Texas Pigtoe Fusconaia askewi G2G3 S3 Ebonyshell Fusconaia ebena G4G5 S3 Round Pearlshell Glebula rotundata G4G5 S4 Pink Mucket Lampsilis abrupta G2 S1 Endangered Endangered Plain Pocketbook Lampsilis cardium G5 S1 Southern Pocketbook Lampsilis ornata G5 S3 Sandbank Pocketbook Lampsilis satura G2 S2 Fatmucket Lampsilis siliquoidea G5 S2 White Heelsplitter Lasmigona complanata G5 S1 Black Sandshell Ligumia recta G4G5 S1 Louisiana Pearlshell Margaritifera hembeli G1 S1 Threatened Threatened Southern Hickorynut Obovaria jacksoniana G2 S1S2 Hickorynut Obovaria olivaria G4 S1 Alabama Hickorynut Obovaria unicolor G3 S1 Mississippi Pigtoe Pleurobema beadleianum G3 S2 Louisiana Pigtoe Pleurobema riddellii G1G2 S1S2 Pyramid Pigtoe Pleurobema rubrum G2G3 S2 Texas Heelsplitter Potamilus amphichaenus G1G2 SH Fat Pocketbook Potamilus capax G2 S1 Endangered Endangered Inflated Heelsplitter Potamilus inflatus G1G2Q S1 Threatened Threatened Ouachita Kidneyshell Ptychobranchus occidentalis G3G4 S1 Rabbitsfoot Quadrula cylindrica G3G4 S1 Threatened Threatened Monkeyface Quadrula metanevra G4 S1 Southern Creekmussel Strophitus subvexus -

Guiding Species Recovery Through Assessment of Spatial And
Guiding Species Recovery through Assessment of Spatial and Temporal Population Genetic Structure of Two Critically Endangered Freshwater Mussel Species (Bivalvia: Unionidae) Jess Walter Jones ( [email protected] ) United States Fish and Wildlife Service Timothy W. Lane Virginia Department of Game and Inland Fisheries N J Eric M. Hallerman Virginia Tech: Virginia Polytechnic Institute and State University Research Article Keywords: Freshwater mussels, Epioblasma brevidens, E. capsaeformis, endangered species, spatial and temporal genetic variation, effective population size, species recovery planning, conservation genetics Posted Date: March 16th, 2021 DOI: https://doi.org/10.21203/rs.3.rs-282423/v1 License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License Page 1/28 Abstract The Cumberlandian Combshell (Epioblasma brevidens) and Oyster Mussel (E. capsaeformis) are critically endangered freshwater mussel species native to the Tennessee and Cumberland River drainages, major tributaries of the Ohio River in the eastern United States. The Clinch River in northeastern Tennessee (TN) and southwestern Virginia (VA) harbors the only remaining stronghold population for either species, containing tens of thousands of individuals per species; however, a few smaller populations are still extant in other rivers. We collected and analyzed genetic data to assist with population restoration and recovery planning for both species. We used an 888 base-pair sequence of the mitochondrial NADH dehydrogenase 1 (ND1) gene and ten nuclear DNA microsatellite loci to assess patterns of genetic differentiation and diversity in populations at small and large spatial scales, and at a 9-year (2004 to 2013) temporal scale, which showed how quickly these populations can diverge from each other in a short time period. -

REPORT FOR: Preliminary Analysis for Identification, Distribution, And
REPORT FOR: Preliminary Analysis for Identification, Distribution, and Conservation Status of Species of Fusconaia and Pleurobema in Arkansas Principle Investigators: Alan D. Christian Department of Biological Sciences, Arkansas State University, P.O. Box 599, State University, Arkansas 72467; [email protected]; Phone: (870)972-3082; Fax: (870)972-2638 John L. Harris Department of Biological Sciences, Arkansas State University, P.O. Box 599, State University, Arkansas 72467 Jeanne Serb Department of Ecology, Evolution, and Organismal Biology, Iowa State University, 251 Bessey Hall, Ames, Iowa 50011 Graduate Research Assistant: David M. Hayes, Department of Environmental Science, P.O. Box 847, State University, Arkansas 72467: [email protected] Kentaro Inoue, Department of Environmental Science, P.O. Box 847, State University, Arkansas 72467: [email protected] Submitted to: William R. Posey Malacologist and Commercial Fisheries Biologist, AGFC P.O. Box 6740 Perrytown, Arkansas 71801 April 2008 EXECUTIVE SUMMARY There are currently 13 species of Fusconaia and 32 species of Pleurobema recognized in the United States and Canada. Twelve species of Pleurobema and two species of Fusconaia are listed as Threatened or Endangered. There are 75 recognized species of Unionidae in Arkansas; however this number may be much higher due to the presence of cryptic species, many which may reside within the Fusconaia /Pleurobema complex. Currently, three species of Fusconaia and three species of Pleurobema are recognized from Arkansas. The true conservation status of species within these genera cannot be determined until the taxonomic identity of populations is confirmed. The purpose of this study was to begin preliminary analysis of the species composition of Fusconaia and Pleurobema in Arkansas and to determine the phylogeographic relationships within these genera through mitochondrial DNA sequencing and conchological analysis. -

Freshwater Mussel Survey of Clinchport, Clinch River, Virginia: Augmentation Monitoring Site: 2006
Freshwater Mussel Survey of Clinchport, Clinch River, Virginia: Augmentation Monitoring Site: 2006 By: Nathan L. Eckert, Joe J. Ferraro, Michael J. Pinder, and Brian T. Watson Virginia Department of Game and Inland Fisheries Wildlife Diversity Division October 28th, 2008 Table of Contents Introduction....................................................................................................................... 4 Objective ............................................................................................................................ 5 Study Area ......................................................................................................................... 6 Methods.............................................................................................................................. 6 Results .............................................................................................................................. 10 Semi-quantitative .................................................................................................. 10 Quantitative........................................................................................................... 11 Qualitative............................................................................................................. 12 Incidental............................................................................................................... 12 Discussion........................................................................................................................ -

Underwater Observation and Habitat Utilization of Three Rare Darters
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Masters Theses Graduate School 5-2010 Underwater observation and habitat utilization of three rare darters (Etheostoma cinereum, Percina burtoni, and Percina williamsi) in the Little River, Blount County, Tennessee Robert Trenton Jett University of Tennessee - Knoxville, [email protected] Follow this and additional works at: https://trace.tennessee.edu/utk_gradthes Part of the Natural Resources and Conservation Commons Recommended Citation Jett, Robert Trenton, "Underwater observation and habitat utilization of three rare darters (Etheostoma cinereum, Percina burtoni, and Percina williamsi) in the Little River, Blount County, Tennessee. " Master's Thesis, University of Tennessee, 2010. https://trace.tennessee.edu/utk_gradthes/636 This Thesis is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Masters Theses by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a thesis written by Robert Trenton Jett entitled "Underwater observation and habitat utilization of three rare darters (Etheostoma cinereum, Percina burtoni, and Percina williamsi) in the Little River, Blount County, Tennessee." I have examined the final electronic copy of this thesis for form and content and recommend that it be accepted in partial fulfillment of the equirr ements for the degree of Master of Science, with a major in Wildlife and Fisheries Science. James L. Wilson, Major Professor We have read this thesis and recommend its acceptance: David A. Etnier, Jason G. -

Species Assessment for Fat Pocketbook
Species Status Assessment Class: Bivalvia Family: Unionidae Scientific Name: Potamilus capax Common Name: Fat pocketbook Species synopsis: Potamilus capax is thought to have been extirpated in New York State for over a century (Strayer and Jirka 1997), and has not been found at historical sites during recent surveys (Mahar & Landry 2013). P. capax is a member of the widely distributed genus Potamilus. P. capax belongs to the subfamily Ambleminae and the tribe Lampsilini, which includes 17 extant and 6 likely extirpated New York species of the genera Actinonaias, Epioblasma, Lampsilis, Leptodea, Ligumia, Obovaria, Potamilus, Ptychobranchus, Toxolasma, Truncilla, and Villosa (Haag 2012; Graf and Cummings 2011). This species is listed as state and federally endangered and is ranked by The Natural Heritage Program as historic in New York and as imperiled throughout its range. It is distributed in the Lower Ohio River system and Mississippi River drainages in Arkansas and Nebraska (Watters et al. 2009), where P. capax abundance is stable, with multiple reproductively viable sites. A current threat to the species is that populations are sporadic and disjunct, with the entire species only inhabiting approximately 20 sites (NatureServe 2013). 1 I. Status a. Current and Legal Protected Status i. Federal ____ Endangered_______________Candidate? ___________ ii. New York _____Endangered__________________________________________________ b. Natural Heritage Program Rank i. Global _____G2- Imperiled_______________________________________________ ii. New York _____SH - Historic_______ Tracked by NYNHP? ______Yes___ Other Rank: U.S. Endangered Species Act (USESA): LE: Listed endangered (1976) IUCN Red List Category: Critically endangered Convention on International Trade in Endangered Species Protection Status (CITES): Appendix I American Fisheries Society Status: Endangered (1993) Status Discussion: The peripheral range (where P. -

September 24, 2018
September 24, 2018 Sent via Federal eRulemaking Portal to: http://www.regulations.gov Docket Nos. FWS-HQ-ES-2018-0006 FWS-HQ-ES-2018-0007 FWS-HQ-ES-2018-0009 Bridget Fahey Chief, Division of Conservation and Classification U.S. Fish and Wildlife Service 5275 Leesburg Pike, MS: ES Falls Church, VA 22041-3808 [email protected] Craig Aubrey Chief, Division of Environmental Review Ecological Services Program U.S. Fish and Wildlife Service 5275 Leesburg Pike, MS: ES Falls Church, VA 22041 [email protected] Samuel D. Rauch, III National Marine Fisheries Service Office of Protected Resources 1315 East-West Highway Silver Spring, MD 20910 [email protected] Re: Proposed Revisions of Endangered Species Act Regulations Dear Mr. Aubrey, Ms. Fahey, and Mr. Rauch: The Southern Environmental Law Center (“SELC”) submits the following comments in opposition to the U.S. Fish and Wildlife Service’s and National Marine Fisheries Service’s proposed revisions to the Endangered Species Act’s implementing regulations.1 We submit these comments on behalf of 57 organizations working to protect the natural resources of the 1 Revision of the Regulations for Prohibitions to Threatened Wildlife and Plants, 83 Fed. Reg. 35,174 (proposed July 25, 2018) (to be codified at 50 C.F.R. pt. 17); Revision of Regulations for Interagency Cooperation, 83 Fed. Reg. 35,178 (proposed July 25, 2018) (to be codified at 50 C.F.R. pt. 402); Revision of the Regulations for Listing Species and Designating Critical Habitat, 83 Fed. Reg. 35,193 (proposed July 25, 2018) (to be codified at 50 C.F.R.