Protection of a Single-Cysteine Redox Switch from Oxidative Destruction: on the Functional Role of Sulfenyl Amide Formation in the Redox-Regulated Enzyme PTP1B
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Norbornene Probes for the Detection of Cysteine Sulfenic Acid in Cells Lisa J
Letters Cite This: ACS Chem. Biol. 2019, 14, 594−598 pubs.acs.org/acschemicalbiology Norbornene Probes for the Detection of Cysteine Sulfenic Acid in Cells Lisa J. Alcock,†,‡ Bruno L. Oliveira,§ Michael J. Deery,∥ Tara L. Pukala,⊥ Michael V. Perkins,† Goncalo̧ J. L. Bernardes,*,§,# and Justin M. Chalker*,†,‡ † Flinders University, College of Science and Engineering, Sturt Road, Bedford Park, South Australia 5042, Australia ‡ Flinders University, Institute for NanoScale Science and Technology, Sturt Road, Bedford Park, South Australia 5042, Australia § Department of Chemistry, University of Cambridge, Lensfield Road, Cambridge CB2 1EW, United Kingdom ∥ Cambridge Centre for Proteomics, Cambridge Systems Biology Centre, Department of Biochemistry, University of Cambridge, Tennis Court Road, Cambridge CB2 1QW, United Kingdom ⊥ The University of Adelaide, School of Physical Sciences, Adelaide, South Australia 5005, Australia # Instituto de Medicina Molecular, Faculdade de Medicina, Universidade de Lisboa, Avenida Professor Egas Moniz, 1649-028, Lisboa, Portugal *S Supporting Information ABSTRACT: Norbornene derivatives were validated as probes for cysteine sulfenic acid on proteins and in live cells. Trapping sulfenic acids with norbornene probes is highly selective and revealed a different reactivity profile than the traditional dimedone reagent. The norbornene probe also revealed a superior chemoselectivity when compared to a commonly used dimedone probe. Together, these results advance the study of cysteine oxidation in biological systems. -

Amide Activation: an Emerging Tool for Chemoselective Synthesis
Featuring work from the research group of Professor As featured in: Nuno Maulide, University of Vienna, Vienna, Austria Amide activation: an emerging tool for chemoselective synthesis Let them stand out of the crowd – Amide activation enables the chemoselective modification of a large variety of molecules while leaving many other functional groups untouched, making it attractive for the synthesis of sophisticated targets. This issue features a review on this emerging field and its application in total synthesis. See Nuno Maulide et al., Chem. Soc. Rev., 2018, 47, 7899. rsc.li/chem-soc-rev Registered charity number: 207890 Chem Soc Rev View Article Online REVIEW ARTICLE View Journal | View Issue Amide activation: an emerging tool for chemoselective synthesis Cite this: Chem. Soc. Rev., 2018, 47,7899 Daniel Kaiser, Adriano Bauer, Miran Lemmerer and Nuno Maulide * It is textbook knowledge that carboxamides benefit from increased stabilisation of the electrophilic carbonyl carbon when compared to other carbonyl and carboxyl derivatives. This results in a considerably reduced reactivity towards nucleophiles. Accordingly, a perception has been developed of amides as significantly less useful functional handles than their ester and acid chloride counterparts. Received 27th April 2018 However, a significant body of research on the selective activation of amides to achieve powerful DOI: 10.1039/c8cs00335a transformations under mild conditions has emerged over the past decades. This review article aims at placing electrophilic amide activation in both a historical context and in that of natural product rsc.li/chem-soc-rev synthesis, highlighting the synthetic applications and the potential of this approach. Creative Commons Attribution 3.0 Unported Licence. -

Technical Notes
1624 Bioconjugate Chem. 2005, 16, 1624−1628 TECHNICAL NOTES Synthesis of Chemical Probes to Map Sulfenic Acid Modifications on Proteins Leslie B. Poole,†* Bu-Bing Zeng,‡,| Sarah A. Knaggs,‡ Mamudu Yakubu,§ and S. Bruce King‡,* Departments of Chemistry and Biochemistry, Wake Forest University, Winston-Salem, North Carolina 27109, and Department of Chemistry, Elizabeth City State University, Elizabeth City, North Carolina 27909 . Received August 23, 2005 Cysteine sulfenic acids in proteins can be identified by their ability to form adducts with dimedone, but this reagent imparts no spectral or affinity tag for subsequent analyses of such tagged proteins. Given its similar reactivity toward cysteine sulfenic acids, 1,3-cyclohexadione was synthetically modified to an alcohol derivative and linked to fluorophores based on isatoic acid and 7-methoxycou- marin. The resulting compounds retain full reactivity and specificity toward cysteine sulfenic acids in proteins, allowing for incorporation of the fluorescent label into the protein and “tagging” it based on its sulfenic acid redox state. Control experiments using dimedone further show the specificity of the reaction of 1,3-diones with protein sulfenic acids in aqueous media. These new compounds provide the basis for an improved method for the detection of protein sulfenic acids. INTRODUCTION Scheme 1 Interest in the identification of cysteine sulfenic acids in proteins by biochemists has grown substantially over the past decade as their biological roles in redox regula- tion and catalysis within an array of cellular proteins have become better defined (1, 2). Despite their impor- tance, only a limited set of tools to identify these species exist, and most of these are only applicable to in vitro studies of pure, isolated proteins (3, 4). -

THIOL OXIDATION a Slippery Slope the Oxidation of Thiols — Molecules RSH Oxidation May Proceed Too Predominates
RESEARCH HIGHLIGHTS Nature Reviews Chemistry | Published online 25 Jan 2017; doi:10.1038/s41570-016-0013 THIOL OXIDATION A slippery slope The oxidation of thiols — molecules RSH oxidation may proceed too predominates. Here, the maximum of the form RSH — can afford quickly for intermediates like RSOH rate constants indicate the order − − − These are many products. From least to most to be spotted and may also afford of reactivity: RSO > RS >> RSO2 . common oxidized, these include disulfides intractable mixtures. Addressing When the reactions are carried out (RSSR), as well as sulfenic (RSOH), the first problem, Chauvin and Pratt in methanol-d , the obtained kinetic reactions, 1 sulfinic (RSO2H) and sulfonic slowed the reactions down by using isotope effect values (kH/kD) are all but have (RSO3H) acids. Such chemistry “very sterically bulky thiols, whose in the range 1.1–1.2, indicating that historically is pervasive in nature, in which corresponding sulfenic acids were no acidic proton is transferred in the been very disulfide bonds between cysteine known to be isolable but were yet rate-determining step. Rather, the residues stabilize protein structures, to be thoroughly explored in terms oxidations involve a specific base- difficult to and where thiols and thiolates often of reactivity”. The second problem catalysed mechanism wherein an study undergo oxidation by H2O2 or O2 in was tackled by modifying the model acid–base equilibrium precedes the order to protect important biological system, 9-mercaptotriptycene, by rate-determining nucleophilic attack − − − structures from damage. Among including a fluorine substituent to of RS , RSO or RSO2 on H2O2. the oxidation products, sulfenic serve as a spectroscopic handle. -

Cysteine Sulfenic Acid As an Intermediate in Disulfide Bond Formation and Nonenzymatic Protein Folding† Douglas S
7748 Biochemistry 2010, 49, 7748–7755 DOI: 10.1021/bi1008694 Cysteine sulfenic Acid as an Intermediate in Disulfide Bond Formation and Nonenzymatic Protein Folding† Douglas S. Rehder and Chad R. Borges* Molecular Biomarkers, The Biodesign Institute at Arizona State University, Tempe, Arizona 85287 Received May 28, 2010; Revised Manuscript Received July 22, 2010 ABSTRACT: As a posttranslational protein modification, cysteine sulfenic acid (Cys-SOH) is well established as an oxidative stress-induced mediator of enzyme function and redox signaling. Data presented herein show that protein Cys-SOH forms spontaneously in air-exposed aqueous solutions of unfolded (disulfide-reduced) protein in the absence of added oxidizing reagents, mediating the oxidative disulfide bond formation process key to in vitro, nonenzymatic protein folding. Molecular oxygen (O2) and trace metals [e.g., copper(II)] are shown to be important reagents in the oxidative refolding process. Cys-SOH is also shown to play a role in spontaneous disulfide-based dimerization of peptide molecules containing free cysteine residues. In total, the data presented expose a chemically ubiquitous role for Cys-SOH in solutions of free cysteine-containing protein exposed to air. In the early 1960s, Anfinsen and colleagues conducted seminal ous oxidants such as hydrogen peroxide are required to build up studies in protein disulfide bond formation and folding, showing significant quantities of Cys-SOH within fully folded proteins. that unfolded (disulfide-reduced) proteins will spontaneously In general, Cys-SOH is a transient intermediate that until the and completely refold, regenerating full protein activity, over a past decade has been difficult to study. It has been understood period of approximately a day in the absence of denaturants that Cys-SOH likely renders the sulfur atom of cysteine electro- (1-6). -

Copper–Sulfenate Complex from Oxidation of a Cavity Mutant of Pseudomonas Aeruginosa Azurin
Copper–sulfenate complex from oxidation of a cavity mutant of Pseudomonas aeruginosa azurin Nathan A. Sierackia,1, Shiliang Tiana,1, Ryan G. Hadtb,1, Jun-Long Zhangc, Julia S. Woertinkb, Mark J. Nilgesa, Furong Suna, Edward I. Solomonb,2, and Yi Lua,2 aDepartment of Chemistry, University of Illinois at Urbana–Champaign, Urbana, IL 61801; bDepartment of Chemistry, Stanford University, Stanford, CA 94305; and cBeijing National Laboratory for Molecular Sciences, State Key Laboratory of Rare Earth Materials Chemistry and Applications, College of Chemistry and Molecular Engineering, Peking University, Beijing 100871, People’s Republic of China Edited by Harry B. Gray, California Institute of Technology, Pasadena, CA, and approved November 27, 2013 (received for review September 3, 2013) Metal–sulfenate centers are known to play important roles in bi- and water. In both enzymes, the sulfenate and sulfinate groups are ology and yet only limited examples are known due to their in- unambiguously required for activity (7, 16). It is commonly postu- stability and high reactivity. Herein we report a copper–sulfenate lated that such oxidized forms of cysteine in a metal site encourage complex characterized in a protein environment, formed at the ac- and stabilize the more oxidized form of redox-active metals, sup- tive site of a cavity mutant of an electron transfer protein, type 1 pressing their potential toxicity in enzymes that do not require the blue copper azurin. Reaction of hydrogen peroxide with Cu(I)– metal center to participate in electron transfer (17, 18). In a more M121G azurin resulted in a species with strong visible absorptions recent example, an iron-coordinated sulfenate was observed when at 350 and 452 nm and a relatively low electron paramagnetic res- a thiolate-containing substrate analog in isopenicillin N-synthase onance gz value of 2.169 in comparison with other normal type 2 was unexpectedly oxidized to a coordinated sulfenato group in copper centers. -
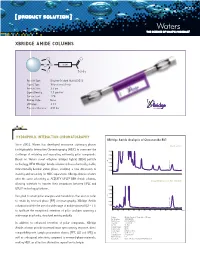
Xbridge Amide Columns
[ PRODUCT SOLUTION ] XBRIDGE AMIDE COLUMNS O O O Si Linker O NH 2 Particle Type: Ethylene Bridged Hybrid [BEH] Ligand Type: Trifunctional Amide Particle Size: 3.5 µm Ligand Density: 7.5 µmol/m2 Carbon Load: 12% Endcap Style: None pH Range: 2-11 Pressure Tolerance: 400 bar HYDROPHILIC INTERacTION CHROMATOGRAPHY XBridge Amide Analysis of Ginsenoside Rb1 Since 2003, Waters has developed innovative stationary phases Orento extract for Hydrophilic Interaction Chromatography [HILIC] to overcome the 0.010 challenge of retaining and separating extremely polar compounds. 0.008 Based on Waters novel ethylene bridged hybrid [BEH] particle 0.006 AU technology, NEW XBridge™ Amide columns utilize a chemically stable, 0.004 0.002 Ginsenoside Rb1 trifunctionally-bonded amide phase, enabling a new dimension in 0.000 stability and versatility for HILIC separations. XBridge Amide columns offer the same selectivity as ACQUITY UPLC® BEH Amide columns, 20 µg/mL Ginsenoside Rb1 standard allowing scientists to transfer their separations between HPLC and 0.010 ® UPLC technology platforms. 0.008 0.006 Designed to retain polar analytes and metabolites that are too polar AU 0.004 to retain by reversed-phase [RP] chromatography, XBridge Amide 0.002 columns facilitate the use of a wide range of mobile phase pH [2 – 11] 0.000 Ginsenoside Rb1 to facilitate the exceptional retention of polar analytes spanning a 0.00. 5 1.01. 5 2.02. 5 3.03. 5 4.04. 5 5.05. 5 6.06. 5 min wide range in polarity, structural moiety and pKa. Column: XBridge Amide, 3.5 µm, 4.6 x 150 mm Part Number: 186004869 Mobile Phase : 80/20 ACN/H2O In addition to enhanced retention of polar compounds, XBridge Flow Rate: 1.4 mL/min Inj. -
In This Handout, All of Our Functional Groups Are Presented As Condensed Line Formulas, 2D and 3D Formulas and with Nomenclature Prefixes and Suffixes (If Present)
In this handout, all of our functional groups are presented as condensed line formulas, 2D and 3D formulas and with nomenclature prefixes and suffixes (if present). Organic names are built on a foundation of alkanes, alkenes and alkynes. Those examples are presented first and you need to know those rules. The strategies can be found in Chapter 4 of our textbook (alkanes: pages 93-98, cycloalkanes 102-104, alkenes: pages 104-110, alkynes: pages 112-113 and combinations of all of them 113-115). After introducing examples of alkanes, alkenes, alkynes and combinations of them, the functional groups are presented in order of priority. A few nomenclature examples are provided for each of the functional groups. Examples of the various functional groups are presented on pages 115-135 in the textbook. Two overview pages are on pages 136-137. Some functional groups have a suffix name when they are the highest priority functional group and a prefix name when they are not the highest priority group, and these are added to the skeletal names with identifying numbers and stereochemistry terms (E and Z for alkenes, R and S for chiral centers and cis and trans for rings). Several low priority functional groups only have a prefix name. A few additional special patterns are shown on pages 98-102. The only way to learn this topic is practice (over and over). The best practice approach is to actually write out the names (on an extra piece of paper or on a white board, and then do it again). The same functional groups are used throughout the entire course. -

UNDERSTANDING SULFUR BASED REDOX BIOLOGY THROUGH ADVANCEMENTS in CHEMICAL BIOLOGY by THOMAS POOLE a Dissertation Submitted to T
UNDERSTANDING SULFUR BASED REDOX BIOLOGY THROUGH ADVANCEMENTS IN CHEMICAL BIOLOGY BY THOMAS POOLE A Dissertation Submitted to the Graduate Faculty of WAKE FOREST UNIVERSITY GRADUATE SCHOOL OF ARTS AND SCIENCES in Partial Fulfillment of the Requirements for the Degree of DOCTOR OF PHILOSOPHY Chemistry May 2017 Winston-Salem, North Carolina Approved By: S. Bruce King, Ph.D., Advisor Leslie B. Poole, Ph.D., Chair Patricia C. Dos Santos, Ph.D. Paul B. Jones, Ph.D. Mark E. Welker, Ph.D. ACKNOWLEDGEMENTS First and foremost I would like to thank professor S. Bruce King. His approach to science makes him an excellent role model that I strive to emulate. He is uniquely masterful at parsing science into important pieces and identifying the gaps and opportunities that others have missed. I thank him for the continued patience and invaluable insight as I’ve progressed through my graduate studies. This work would not have been possible without his insight and guiding suggestions. I would like to thank our collaborators who have proven invaluable in their support. Cristina Furdui has provided the knowledge, context, and biochemical support that allowed our work to be published in esteemed journals. Leslie Poole has provided the insight into redox biology and guidance towards appropriate experiments. I thank my committee members, Mark Welker and Patricia Dos Santos who have guided my graduate work from the beginning. Professor Dos Santos provided the biological perspective to evaluate my redox biology work. In addition, she helped guide the biochemical understanding required to legitimize my independent proposal. Professor Welker provided the scrutinizing eye for my early computational work and suggested validating the computational work with experimental data. -

United States Patent Office Patented Nov
3,285,929 United States Patent Office Patented Nov. 15, 1966 1. 2 3,285,929 and R represents an organic radical or a hydrogen radi FUNGCDAL COMPOUNDS CONTAINING cal. THE NSCFC-GROUPS Suitable imide compounds of the indicated type are Erich Kauke, Cologne-Fliétard, Engelbert Kuhle and illustrated by Formula II: Ferdinand Grewe, Cologne-Stammheim, and Helmut Kaspers and Richard Wegler, Leverkusen, Germany, (II) X assignors to Farbenfabriken Bayer Aktiengesellschaft, R^ Yscrol, Leverkusen, Germany, a corporation of Germany No Drawing. Filed Oct. 31, 1961, Ser. No. 148,845 Y Claims priority, application Germany, Nov. 3, 1960, in which R represents one or more organic residues and F 32,468; Nov. 25, 1960, F 32,620 0. X is part of an acyl group, such as for example 8 Claims. (C. 260-301) O O The present invention relates to new and useful im provements in fungicidal preparations and more particu -- and -C- larly to improved fungicides. This invention also relates O to methods of protecting organic material subject to at 15 The organic residue may be aliphatic, aromatic, ali tack by low orders of organisms. This invention further cyclic, heterocyclic, and their substituted derivatives, and relates to the synthesis of new chemical compounds, may be attached furthermore to the above said group namely N-thiomonofluoro-dichloromethyl imides of the also by heteroatoms such as nitrogen, sulfur or oxygen. following general formula The novel compounds of this invention may be pre 20 pared in general by the reaction of dichloro-monofluoro methyl-sulf-enylchloride (CISCFC), with the corre sponding imide or the metal salt of this compound. -

Detection of Atmospheric Gaseous Amines and Amides by a High-Resolution Time-Of-flight Chemical Ionization Mass Spectrometer with Protonated Ethanol Reagent Ions
Atmos. Chem. Phys., 16, 14527–14543, 2016 www.atmos-chem-phys.net/16/14527/2016/ doi:10.5194/acp-16-14527-2016 © Author(s) 2016. CC Attribution 3.0 License. Detection of atmospheric gaseous amines and amides by a high-resolution time-of-flight chemical ionization mass spectrometer with protonated ethanol reagent ions Lei Yao1, Ming-Yi Wang1,a, Xin-Ke Wang1, Yi-Jun Liu1,b, Hang-Fei Chen1, Jun Zheng2, Wei Nie3,4, Ai-Jun Ding3,4, Fu-Hai Geng5, Dong-Fang Wang6, Jian-Min Chen1, Douglas R. Worsnop7, and Lin Wang1,4 1Shanghai Key Laboratory of Atmospheric Particle Pollution and Prevention (LAP3), Department of Environmental Science & Engineering, Fudan University, Shanghai 200433, China 2Jiangsu Key Laboratory of Atmospheric Environment Monitoring and Pollution Control, Nanjing University of Information Science & Technology, Nanjing 210044, China 3Joint International Research Laboratory of Atmospheric and Earth System Sciences, School of Atmospheric Science, Nanjing University, Nanjing 210023, China 4Collaborative Innovation Center of Climate Change, Nanjing 210023, China 5Shanghai Meteorology Bureau, Shanghai 200135, China 6Shanghai Environmental Monitoring Center, Shanghai 200030, China 7Aerodyne Research, Billerica, MA 01821, USA anow at: Center for Atmospheric Particle Studies, Carnegie Mellon University, Pittsburgh, PA 15213, USA bnow at: Pratt School of Engineering, Duke University, Durham, NC 27705, USA Correspondence to: Lin Wang([email protected]) Received: 7 June 2016 – Published in Atmos. Chem. Phys. Discuss.: 22 June 2016 Revised: 15 October 2016 – Accepted: 3 November 2016 – Published: 23 November 2016 Abstract. Amines and amides are important atmospheric concentrations of amines ranged from a few parts per trillion organic-nitrogen compounds but high time resolution, highly by volume to hundreds of parts per trillion by volume, con- sensitive, and simultaneous ambient measurements of these centrations of amides varied from tens of parts per trillion by species are rather sparse. -

Conferring Specificity in Redox Pathways by Enzymatic Thiol/Disulfide Exchange Reactions
Free Radical Research ISSN: 1071-5762 (Print) 1029-2470 (Online) Journal homepage: https://www.tandfonline.com/loi/ifra20 Conferring specificity in redox pathways by enzymatic thiol/disulfide exchange reactions Luis Eduardo S. Netto, Marcos Antonio de Oliveira, Carlos A. Tairum & José Freire da Silva Neto To cite this article: Luis Eduardo S. Netto, Marcos Antonio de Oliveira, Carlos A. Tairum & José Freire da Silva Neto (2016) Conferring specificity in redox pathways by enzymatic thiol/disulfide exchange reactions, Free Radical Research, 50:2, 206-245, DOI: 10.3109/10715762.2015.1120864 To link to this article: https://doi.org/10.3109/10715762.2015.1120864 Accepted author version posted online: 16 Nov 2015. Published online: 08 Jan 2016. Submit your article to this journal Article views: 727 View Crossmark data Citing articles: 24 View citing articles Full Terms & Conditions of access and use can be found at https://www.tandfonline.com/action/journalInformation?journalCode=ifra20 FREE RADICAL RESEARCH, 2016 VOL. 50, NO. 2, 206–245 http://dx.doi.org/10.3109/10715762.2015.1120864 REVIEW ARTICLE Conferring specificity in redox pathways by enzymatic thiol/disulfide exchange reactions Luis Eduardo S. Nettoa, Marcos Antonio de Oliveirab, Carlos A. Tairumb and Jose´ Freire da Silva Netoc aDepartamento de Gene´tica e Biologia Evolutiva, Instituto de Biocieˆncias, Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil; bDepartamento de Biologia, Universidade Estadual Paulista Ju´lio de Mesquita Filho, Campus do Litoral Paulista Sa˜o Vicente, Brazil; cDepartamento de Biologia Celular e Molecular e Bioagentes Patogeˆnicos, Faculdade de Medicina de Ribeira˜o Preto, Universidade de Sa˜o Paulo, Ribeira˜o Preto, Sa˜o Paulo, Brazil ABSTRACT ARTICLE HISTORY Thiol–disulfide exchange reactions are highly reversible, displaying nucleophilic substitutions Received 20 August 2015 mechanism (SN2 type).