The Influence of Optical Distortions and Transverse Chromatic Aberrations on Motion Parallax and Stereopsis in Natural and Artificial Environments
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
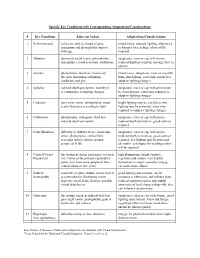
Specific Eye Conditions with Corresponding Adaptations/Considerations
Specific Eye Conditions with Corresponding Adaptations/Considerations # Eye Condition Effect on Vision Adaptations/Considerations 1 Achromotopsia colors are seen as shades of grey, tinted lenses, reduced lighting, alternative nystagmus and photophobia improve techniques for teaching colors will be with age required 2 Albinism decreased visual acuity, photophobia, sunglasses, visor or cap with a brim, nystagmus, central scotomas, strabismus reduced depth perception, moving close to objects 3 Aniridia photophobia, field loss, vision may tinted lenses, sunglasses, visor or cap with fluctuate depending on lighting brim, dim lighting, extra time required to conditions and glare adapt to lighting changes 4 Aphakia reduced depth perception, inability to sunglasses, visor or cap with a brim may accommodate to lighting changes be worn indoors, extra time required to adapt to lighting changes 5 Cataracts poor color vision, photophobia, visual bright lighting may be a problem, low acuity fluctuates according to light lighting may be preferred, extra time required to adapt to lighting changes 6 Colobomas photophobia, nystagmus, field loss, sunglasses, visor or cap with a brim, reduced depth perception reduced depth perception, good contrast required 7 Color Blindness difficulty or inability to see colors and sunglasses, visor or cap with a brim, detail, photophobia, central field reduced depth perception, good contrast scotomas (spotty vision), normal required, low lighting may be preferred, peripheral fields alternative techniques for teaching colors -

Spacespex™ Anaglyph—The Only Way to Bring 3Dtv to the Masses
SPACESPEX™ ANAGLYPH—THE ONLY WAY TO BRING 3DTV TO THE MASSES By Michael Starks © M. Starks 2009 May be reproduced provided nothing is added, omitted or changed-- including this copyright notice. SpaceSpex™ is the name I applied to my versions of the orange/blue anaglyph technique in 1993. In fact the Gang Li/ColorCode and some models of SpaceSpex use amber or orange/brown rather than yellow, but they are on a continuum. Like all the bicolor anaglyph methods it is compatible with all video equipment and displays and I think it’s the best of the methods using inexpensive paper glasses with colored lenses. Until someone comes up with a way to put hundreds of millions of new 3D TV’s in homes which can use polarized glasses or LCD shutter glasses, anaglyph is going to be the only way for mass distribution of full color high quality 3D over cable, satellite, the web or on DVD. However the solution I have proposed for Set Top Boxes, PC’s, TV sets and DVD players for the last 20 years is to have user controls, so those with display hardware that permits polarized or shutter glasses or even autostereo viewing or who want 2D can make that choice from the single 3D video file. This is the method of the TDVision codec, Next3D, and of Peter Wimmer’s famous StereoScopic Player (a new version due end of 2009), (all of which should appear in hardware soon) and probably the best stereoplayer of all in Masuji Suto’s StereoMovie Maker, and is being incorporated in most well known software DVD and media players. -

Teacher Guide
Neuroscience for Kids http://faculty.washington.edu/chudler/neurok.html. Our Sense of Sight: Part 2. Perceiving motion, form, and depth Visual Puzzles Featuring a “Class Experiment” and “Try Your Own Experiment” Teacher Guide WHAT STUDENTS WILL DO · TEST their depth perception using one eye and then two · CALCULATE the class averages for the test perception tests · DISCUSS the functions of depth perception · DEFINE binocular vision · IDENTIFY monocular cues for depth · DESIGN and CONDUCT further experiments on visual perception, for example: · TEST people’s ability to interpret visual illusions · CONSTRUCT and test new visual illusions · DEVISE a “minimum difference test” for visual attention 1 SETTING UP THE LAB Supplies For the Introductory Activity Two pencils or pens for each student For the Class Experiment For each group of four (or other number) students: Measuring tools (cloth tape or meter sticks) Plastic cups, beakers or other sturdy containers Small objects such as clothespins, small legos, paper clips For “Try Your Own Experiment!” Visual illusion figures, found at the end of this Teacher Guide Paper and markers or pens Rulers Other Preparations · For the Class Experiment and Do Your Own Experiment, students can write results on a plain sheet of paper. · Construct a chart on the board where data can be entered for class discussion. · Decide the size of the student groups; three is a convenient number for these experiments—a Subject, a Tester, and a Recorder. Depending on materials available, four or five students can comprise a group. · For “Try Your Own Experiment!,” prepare materials in the Supply list and put them out on an “Explore” table. -

Visual Secret Sharing Scheme with Autostereogram*
Visual Secret Sharing Scheme with Autostereogram* Feng Yi, Daoshun Wang** and Yiqi Dai Department of Computer Science and Technology, Tsinghua University, Beijing, 100084, China Abstract. Visual secret sharing scheme (VSSS) is a secret sharing method which decodes the secret by using the contrast ability of the human visual system. Autostereogram is a single two dimensional (2D) image which becomes a virtual three dimensional (3D) image when viewed with proper eye convergence or divergence. Combing the two technologies via human vision, this paper presents a new visual secret sharing scheme called (k, n)-VSSS with autostereogram. In the scheme, each of the shares is an autostereogram. Stacking any k shares, the secret image is recovered visually without any equipment, but no secret information is obtained with less than k shares. Keywords: visual secret sharing scheme; visual cryptography; autostereogram 1. Introduction In 1979, Blakely and Shamir[1-2] independently invented a secret sharing scheme to construct robust key management scheme. A secret sharing scheme is a method of sharing a secret among a group of participants. In 1994, Naor and Shamir[3] firstly introduced visual secret sharing * Supported by National Natural Science Foundation of China (No. 90304014) ** E-mail address: [email protected] (D.S.Wang) 1 scheme in Eurocrypt’94’’ and constructed (k, n)-threshold visual secret sharing scheme which conceals the original data in n images called shares. The original data can be recovered from the overlap of any at least k shares through the human vision without any knowledge of cryptography or cryptographic computations. With the development of the field, Droste[4] provided a new (k, n)-VSSS algorithm and introduced a model to construct the (n, n)-combinational threshold scheme. -

Stereoscopic Therapy: Fun Or Remedy?
STEREOSCOPIC THERAPY: FUN OR REMEDY? SARA RAPOSO Abstract (INDEPENDENT SCHOLAR , PORTUGAL ) Once the material of playful gatherings, stereoscop ic photographs of cities, the moon, landscapes and fashion scenes are now cherished collectors’ items that keep on inspiring new generations of enthusiasts. Nevertheless, for a stereoblind observer, a stereoscopic photograph will merely be two similar images placed side by side. The perspective created by stereoscop ic fusion can only be experienced by those who have binocular vision, or stereopsis. There are several caus es of a lack of stereopsis. They include eye disorders such as strabismus with double vision. Interestingly, stereoscopy can be used as a therapy for that con dition. This paper approaches this kind of therapy through the exploration of North American collections of stereoscopic charts that were used for diagnosis and training purposes until recently. Keywords. binocular vision; strabismus; amblyopia; ste- reoscopic therapy; optometry. 48 1. Binocular vision and stone (18021875), which “seem to have access to the visual system at the same stereopsis escaped the attention of every philos time and form a unitary visual impres opher and artist” allowed the invention sion. According to the suppression the Vision and the process of forming im of a “simple instrument” (Wheatstone, ory, both similar and dissimilar images ages, is an issue that has challenged 1838): the stereoscope. Using pictures from the two eyes engage in alternat the most curious minds from the time of as a tool for his study (Figure 1) and in ing suppression at a low level of visual Aristotle and Euclid to the present day. -

British Orthoptic Journal Volume 1, 1939
British Orthoptic Journal Volume 1, 1939 Loss of Central Fixation 15-19 I.Yoxall Some Observations on Partial Occlusion in Accommodative Squints 20- 22 E.Pemberton A few samples of Traumatic Heterophoria 23-27 S.Mayou Operative Impressions in Orthoptic Training 28-33 OM Duthie Summary of routine treatment given at the Manchester Royal 34-37 Eye Hospital E.Stringer Voluntary Diplopia 38-43 S.Mayou Paralysis of External Rectus; Treatment of Muscle Grafting 44-45 P.Jameson Evans Some Observations on Squint Operations 46-49 Dr Gordon Napier Suitability of cases for Orthoptic Training 50-53 CH.Bamford Occlusion 54-57 K.Bastow Some Recent Methods used in an Attempt to shorten Orthoptic treatment 58-62 S.Jackson History of Orthoptic treatment 63-65 CL Gimblett Occasional Divergent Squint 66 S.Jackson Unusual cases of divergent squint treated at the Manchester Royal Eye 67 Hospital E.Stringer Divergent Strabismus and its treatments 68-69 K.Bastow Divergent Squint 70 J.Strickland Approach to the Phorias 71-104 Wing Commander Livingstone Certain aspects of the Evolution of the eye 105 I.Mann BOJ Volume 2, 1944 Some Observations on accommodative squint 13-15 M.Parsons Some Observations on experimental work on the relation of squint 16-20 to emotional disturbances carried out at the Oxford Eye Hospital B.Hare Our Failures 21-24 E.Stringer Graded Squint Operations 25-32 J.Foster, EC Pemberton, SS Freedman Prognosis of postoperative Diplopia in adult Squints 33-35 EC Pemberton Some notes on treatment of abnormal retinal correspondence 36-37 B.Hare Convergence -

Neural Correlates of Convergence Eye Movements in Convergence Insufficiency Patients Vs
Neural Correlates of Convergence Eye Movements in Convergence Insufficiency Patients vs. Normal Binocular Vision Controls: An fMRI Study A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in Biomedical Engineering by Chirag B. Limbachia B.S.B.M.E., Wright State University, 2014 2015 Wright State University Wright State University GRADUATE SCHOOL January 18, 2016 I HEREBY RECOMMEND THAT THE THESIS PREPARED UNDER MY SUPER- VISION BY Chirag B. Limbachia ENTITLED Neural Correlates of Convergence Eye Movements in Convergence Insufficiency Patients vs. Normal Binocular Vision Controls: An fMRI Study BE ACCEPTED IN PARTIAL FULFILLMENT OF THE REQUIRE- MENTS FOR THE DEGREE OF Master of Science in Biomedical Engineering. Nasser H. Kashou Thesis Director Jaime E. Ramirez-Vick, Ph.D., Chair, Department of Biomedical, Industrial, and Human Factors Engineering Committee on Final Examination Nasser H. Kashou, Ph.D. Marjean T. Kulp, O.D., M.S., F.A.A.O. Subhashini Ganapathy, Ph.D. Robert E.W. Fyffe, Ph.D. Vice President for Research and Dean of the Graduate School ABSTRACT Limbachia, Chirag. M.S.B.M.E., Department of Biomedical, Industrial, and Human Factor En- gineering, Wright State University, 2015. Neural Correlates of Convergence Eye Movements in Convergence Insufficiency Patients vs. Normal Binocular Vision Controls: An fMRI Study. Convergence Insufficiency is a binocular vision disorder, characterized by reduced abil- ity of performing convergence eye movements. Absence of convergence causes, eye strain, blurred vision, doubled vision, headaches, and difficulty reading due frequent loss of place. These symptoms commonly occur during near work. The purpose of this study was to quantify neural correlates associated with convergence eye movements in convergence in- sufficient (CI) patients vs. -

Relative Importance of Binocular Disparity and Motion Parallax for Depth Estimation: a Computer Vision Approach
remote sensing Article Relative Importance of Binocular Disparity and Motion Parallax for Depth Estimation: A Computer Vision Approach Mostafa Mansour 1,2 , Pavel Davidson 1,* , Oleg Stepanov 2 and Robert Piché 1 1 Faculty of Information Technology and Communication Sciences, Tampere University, 33720 Tampere, Finland 2 Department of Information and Navigation Systems, ITMO University, 197101 St. Petersburg, Russia * Correspondence: pavel.davidson@tuni.fi Received: 4 July 2019; Accepted: 20 August 2019; Published: 23 August 2019 Abstract: Binocular disparity and motion parallax are the most important cues for depth estimation in human and computer vision. Here, we present an experimental study to evaluate the accuracy of these two cues in depth estimation to stationary objects in a static environment. Depth estimation via binocular disparity is most commonly implemented using stereo vision, which uses images from two or more cameras to triangulate and estimate distances. We use a commercial stereo camera mounted on a wheeled robot to create a depth map of the environment. The sequence of images obtained by one of these two cameras as well as the camera motion parameters serve as the input to our motion parallax-based depth estimation algorithm. The measured camera motion parameters include translational and angular velocities. Reference distance to the tracked features is provided by a LiDAR. Overall, our results show that at short distances stereo vision is more accurate, but at large distances the combination of parallax and camera motion provide better depth estimation. Therefore, by combining the two cues, one obtains depth estimation with greater range than is possible using either cue individually. -

Care of the Patient with Accommodative and Vergence Dysfunction
OPTOMETRIC CLINICAL PRACTICE GUIDELINE Care of the Patient with Accommodative and Vergence Dysfunction OPTOMETRY: THE PRIMARY EYE CARE PROFESSION Doctors of optometry are independent primary health care providers who examine, diagnose, treat, and manage diseases and disorders of the visual system, the eye, and associated structures as well as diagnose related systemic conditions. Optometrists provide more than two-thirds of the primary eye care services in the United States. They are more widely distributed geographically than other eye care providers and are readily accessible for the delivery of eye and vision care services. There are approximately 36,000 full-time-equivalent doctors of optometry currently in practice in the United States. Optometrists practice in more than 6,500 communities across the United States, serving as the sole primary eye care providers in more than 3,500 communities. The mission of the profession of optometry is to fulfill the vision and eye care needs of the public through clinical care, research, and education, all of which enhance the quality of life. OPTOMETRIC CLINICAL PRACTICE GUIDELINE CARE OF THE PATIENT WITH ACCOMMODATIVE AND VERGENCE DYSFUNCTION Reference Guide for Clinicians Prepared by the American Optometric Association Consensus Panel on Care of the Patient with Accommodative and Vergence Dysfunction: Jeffrey S. Cooper, M.S., O.D., Principal Author Carole R. Burns, O.D. Susan A. Cotter, O.D. Kent M. Daum, O.D., Ph.D. John R. Griffin, M.S., O.D. Mitchell M. Scheiman, O.D. Revised by: Jeffrey S. Cooper, M.S., O.D. December 2010 Reviewed by the AOA Clinical Guidelines Coordinating Committee: David A. -

Chromostereo.Pdf
ChromoStereoscopic Rendering for Trichromatic Displays Le¨ıla Schemali1;2 Elmar Eisemann3 1Telecom ParisTech CNRS LTCI 2XtremViz 3Delft University of Technology Figure 1: ChromaDepth R glasses act like a prism that disperses incoming light and induces a differing depth perception for different light wavelengths. As most displays are limited to mixing three primaries (RGB), the depth effect can be significantly reduced, when using the usual mapping of depth to hue. Our red to white to blue mapping and shading cues achieve a significant improvement. Abstract The chromostereopsis phenomenom leads to a differing depth per- ception of different color hues, e.g., red is perceived slightly in front of blue. In chromostereoscopic rendering 2D images are produced that encode depth in color. While the natural chromostereopsis of our human visual system is rather low, it can be enhanced via ChromaDepth R glasses, which induce chromatic aberrations in one Figure 2: Chromostereopsis can be due to: (a) longitunal chro- eye by refracting light of different wavelengths differently, hereby matic aberration, focus of blue shifts forward with respect to red, offsetting the projected position slightly in one eye. Although, it or (b) transverse chromatic aberration, blue shifts further toward might seem natural to map depth linearly to hue, which was also the the nasal part of the retina than red. (c) Shift in position leads to a basis of previous solutions, we demonstrate that such a mapping re- depth impression. duces the stereoscopic effect when using standard trichromatic dis- plays or printing systems. We propose an algorithm, which enables an improved stereoscopic experience with reduced artifacts. -

Two Eyes See More Than One Human Beings Have Two Eyes Located About 6 Cm (About 2.4 In.) Apart
ivi act ty 2 TTwowo EyesEyes SeeSee MoreMore ThanThan OneOne OBJECTIVES 1 straw 1 card, index (cut in half widthwise) Students discover how having two eyes helps us see in three dimensions and 3 pennies* increases our field of vision. 1 ruler, metric* The students For the class discover that each eye sees objects from a 1 roll string slightly different viewpoint to give us depth 1 roll tape, masking perception 1 pair scissors* observe that depth perception decreases *provided by the teacher with the use of just one eye measure their field of vision observe that their field of vision decreases PREPARATION with the use of just one eye Session I 1 Make a copy of Activity Sheet 2, Part A, for each student. SCHEDULE 2 Each team of two will need a metric ruler, Session I About 30 minutes three paper cups, and three pennies. Session II About 40 minutes Students will either close or cover their eyes, or you may use blindfolds. (Students should use their own blindfold—a bandanna or long strip VOCABULARY of cloth brought from home and stored in their science journals for use—in this depth perception and other activities.) field of vision peripheral vision Session II 1 Make a copy of Activity Sheet 2, Part B, for each student. MATERIALS 2 For each team, cut a length of string 50 cm (about 20 in.) long. Cut enough index For each student cards in half (widthwise) to give each team 1 Activity Sheet 2, Parts A and B half a card. Snip the corners of the cards to eliminate sharp edges. -

Binocular Vision
BINOCULAR VISION Rahul Bhola, MD Pediatric Ophthalmology Fellow The University of Iowa Department of Ophthalmology & Visual Sciences posted Jan. 18, 2006, updated Jan. 23, 2006 Binocular vision is one of the hallmarks of the human race that has bestowed on it the supremacy in the hierarchy of the animal kingdom. It is an asset with normal alignment of the two eyes, but becomes a liability when the alignment is lost. Binocular Single Vision may be defined as the state of simultaneous vision, which is achieved by the coordinated use of both eyes, so that separate and slightly dissimilar images arising in each eye are appreciated as a single image by the process of fusion. Thus binocular vision implies fusion, the blending of sight from the two eyes to form a single percept. Binocular Single Vision can be: 1. Normal – Binocular Single vision can be classified as normal when it is bifoveal and there is no manifest deviation. 2. Anomalous - Binocular Single vision is anomalous when the images of the fixated object are projected from the fovea of one eye and an extrafoveal area of the other eye i.e. when the visual direction of the retinal elements has changed. A small manifest strabismus is therefore always present in anomalous Binocular Single vision. Normal Binocular Single vision requires: 1. Clear Visual Axis leading to a reasonably clear vision in both eyes 2. The ability of the retino-cortical elements to function in association with each other to promote the fusion of two slightly dissimilar images i.e. Sensory fusion. 3. The precise co-ordination of the two eyes for all direction of gazes, so that corresponding retino-cortical element are placed in a position to deal with two images i.e.