Southern Caribbean Azooxanthellate Coral Communities Off Colombia
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Acanthochitona Pygmea (Pilsbry, 1893)
Lista de especies del phylum Mollusca registradas para el Caribe colombiano Abra aequalis (Say, 1822) Abra longicallis (Sacchi, 1837) Abralia veranyi (Rüppell, 1844) Acanthochitona pygmea (Pilsbry, 1893) Acanthochitona rodea (Pilsbry, 1893) Acanthochitona spiculosa (Reeve, 1847) Acanthochitona venezuelana Lyons, 1888 Acanthopleura granulata (Gmelin, 1791) Acar domingensis (Lamarck, 1819) Acesta colombiana (H.E. Vokes, 1970) Acmaea antillarum (Sowerby, 1831) Acmaea leucopleura (Gmelin, 1791) Acmaea pustulata (Helbling, 1779) Acteocina candei (d'Orbigny, 1842) Acteocina recta (d'Orbigny, 1841) Acteon danaida Dall, 1881 Acteon punctostriatus (C.B. Adams, 1840) Actinotrophon actinophorus (Dall, 1889) Adrana gloriosa (A. Adams, 1855) Adrana patagonica (d'Orbigny, 1846) Adrana scaphoides Rehder, 1939 Adrana tellinoides (Sowerby, 1823) Aesopus obesus (Hinds, 1843) Aesopus stearnsi (Tryon, 1883) Agathotoma badia (Reeve, 1846) Agathotoma candidísima (C.B. Adams, 1850) Agatrix smithi (Dall, 1888) Agladrillia rhodochroa (Dautzenberg, 1900) Akera bayeri Marcus y Marcus, 1967 Alaba incerta (d'Orbigny, 1842) Alvania avernas (C.B. Adams, 1850) Alvania auberiana (d'Orbigny, 1842) Alvania colombiana Romer y Moore, 1988 Amaea mitchelli (Dall, 1889) Amaea retiñera (Dall, 1889) Americardia media (Linné, 1758) Amusium laurenti (Gmelin, 1791) Amusium payraceum (Gabb, 1873) Amygdalum politum (Verrill y Smith, 1880) Amygdalum sagittatum Rehder, 1934 Anachis cf. fraudans Jung, 1969 Anachis coseli Díaz y Mittnacht, 1991 Anachis hotessieriana (d'Orbigny, 1842) -

ABSTRACT Title of Dissertation: PATTERNS IN
ABSTRACT Title of Dissertation: PATTERNS IN DIVERSITY AND DISTRIBUTION OF BENTHIC MOLLUSCS ALONG A DEPTH GRADIENT IN THE BAHAMAS Michael Joseph Dowgiallo, Doctor of Philosophy, 2004 Dissertation directed by: Professor Marjorie L. Reaka-Kudla Department of Biology, UMCP Species richness and abundance of benthic bivalve and gastropod molluscs was determined over a depth gradient of 5 - 244 m at Lee Stocking Island, Bahamas by deploying replicate benthic collectors at five sites at 5 m, 14 m, 46 m, 153 m, and 244 m for six months beginning in December 1993. A total of 773 individual molluscs comprising at least 72 taxa were retrieved from the collectors. Analysis of the molluscan fauna that colonized the collectors showed overwhelmingly higher abundance and diversity at the 5 m, 14 m, and 46 m sites as compared to the deeper sites at 153 m and 244 m. Irradiance, temperature, and habitat heterogeneity all declined with depth, coincident with declines in the abundance and diversity of the molluscs. Herbivorous modes of feeding predominated (52%) and carnivorous modes of feeding were common (44%) over the range of depths studied at Lee Stocking Island, but mode of feeding did not change significantly over depth. One bivalve and one gastropod species showed a significant decline in body size with increasing depth. Analysis of data for 960 species of gastropod molluscs from the Western Atlantic Gastropod Database of the Academy of Natural Sciences (ANS) that have ranges including the Bahamas showed a positive correlation between body size of species of gastropods and their geographic ranges. There was also a positive correlation between depth range and the size of the geographic range. -

The Lower Pliocene Gastropods of Le Pigeon Blanc (Loire- Atlantique, Northwest France). Part 5* – Neogastropoda (Conoidea) and Heterobranchia (Fine)
Cainozoic Research, 18(2), pp. 89-176, December 2018 89 The lower Pliocene gastropods of Le Pigeon Blanc (Loire- Atlantique, northwest France). Part 5* – Neogastropoda (Conoidea) and Heterobranchia (fine) 1 2 3,4 Luc Ceulemans , Frank Van Dingenen & Bernard M. Landau 1 Avenue Général Naessens de Loncin 1, B-1330 Rixensart, Belgium; email: [email protected] 2 Cambeenboslaan A 11, B-2960 Brecht, Belgium; email: [email protected] 3 Naturalis Biodiversity Center, P.O. Box 9517, 2300 RA Leiden, Netherlands; Instituto Dom Luiz da Universidade de Lisboa, Campo Grande, 1749-016 Lisboa, Portugal; and International Health Centres, Av. Infante de Henrique 7, Areias São João, P-8200 Albufeira, Portugal; email: [email protected] 4 Corresponding author Received 25 February 2017, revised version accepted 7 July 2018 In this final paper reviewing the Zanclean lower Pliocene assemblage of Le Pigeon Blanc, Loire-Atlantique department, France, which we consider the ‘type’ locality for Assemblage III of Van Dingenen et al. (2015), we cover the Conoidea and the Heterobranchia. Fifty-nine species are recorded, of which 14 are new: Asthenotoma lanceolata nov. sp., Aphanitoma marqueti nov. sp., Clathurella pierreaimei nov. sp., Clavatula helwerdae nov. sp., Haedropleura fratemcontii nov. sp., Bela falbalae nov. sp., Raphitoma georgesi nov. sp., Raphitoma landreauensis nov. sp., Raphitoma palumbina nov. sp., Raphitoma turtaudierei nov. sp., Raphitoma vercingetorixi nov. sp., Raphitoma pseudoconcinna nov. sp., Adelphotectonica bieleri nov. sp., and Ondina asterixi nov. sp. One new name is erected: Genota maximei nov. nom. is proposed for Pleurotoma insignis Millet, non Edwards, 1861. Actaeonidea achatina Sacco, 1896 is considered a junior subjective synonym of Rictaxis tornatus (Millet, 1854). -

NEAT Mollusca
NEAT (North East Atlantic Taxa): Scandinavian marine Mollusca Check-List compiled at TMBL (Tjärnö Marine Biological Laboratory) by: Hans G. Hansson 1994-02-02 / small revisions until February 1997, when it was published on Internet as a pdf file and then republished August 1998.. Citation suggested: Hansson, H.G. (Comp.), NEAT (North East Atlantic Taxa): Scandinavian marine Mollusca Check-List. Internet Ed., Aug. 1998. [http://www.tmbl.gu.se]. Denotations: (™) = Genotype @ = Associated to * = General note PHYLUM, CLASSIS, SUBCLASSIS, SUPERORDO, ORDO, SUBORDO, INFRAORDO, Superfamilia, Familia, Subfamilia, Genus & species N.B.: This is one of several preliminary check-lists, covering S Scandinavian marine animal (and partly marine protoctistan) taxa. Some financial support from (or via) NKMB (Nordiskt Kollegium för Marin Biologi), during the last years of the existence of this organization (until 1993), is thankfully acknowledged. The primary purpose of these checklists is to faciliate for everyone, trying to identify organisms from the area, to know which species that earlier have been encountered there, or in neighbouring areas. A secondary purpose is to faciliate for non-experts to put as correct names as possible on organisms, including names of authors and years of description. So far these checklists are very preliminary. Due to restricted access to literature there are (some known, and probably many unknown) omissions in the lists. Certainly also several errors may be found, especially regarding taxa like Plathelminthes and Nematoda, where the experience of the compiler is very rudimentary, or. e.g. Porifera, where, at least in certain families, taxonomic confusion seems to prevail. This is very much a small modernization of T. -
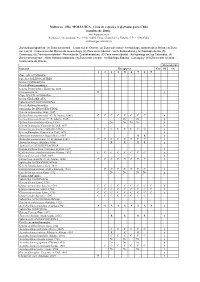
Moluscos - Filo MOLLUSCA
Moluscos - Filo MOLLUSCA. Lista de especies registradas para Cuba (octubre de 2006). José Espinosa Sáez Instituto de Oceanología, Ave 1ª No. 18406, Playa, Ciudad de La Habana, C.P. 11200, Cuba [email protected] Zonas biogeográficas: (1) Zona suroriental – Costa sur de Oriente, (2) Zona surcentral - Archipiélago Jardines de la Reina, (3) Zona sur central - Costa al sur del Macizo de Guamuhaya, (4) Zona suroccidental - Golfo de Batabanó y Archipiélago de los, (5) Canarreos, (6) Zona suroccidental - Península de Guanahacabibes, (7) Zona noroccidental - Archipiélago de Los Colorados, (8) Zona noroccidental - Norte Habana-Matanzas, (9) Zona norte-central - Archipiélago Sabana - Camagüey, (10) Zona norte-oriental - Costa norte de Oriente Abreviaturas Especies Bioegiones Cu Pl Oc 1 2 3 4 5 6 7 8 9 Clase APLACOPHORA Subclase SOLENOGASTRES Orden CAVIBELONIA Familia Proneomeniidae Género Proneomenia Hubrecht, 1880 Proneomenia sp . R x Clase POLYPLACOPHORA Orden NEOLORICATA Suborden ISCHNOCHITONINA Familia Ischnochitonidae Subfamilia ISCHNOCHITONINAE Género Ischnochiton Gray, 1847 Ischnochiton erythronotus (C. B. Adams, 1845) C C C C C C C C x Ischnochiton papillosus (C. B. Adams, 1845) Nc Nc x Ischnochiton striolatus (Gray, 1828) Nc Nc Nc Nc x Género Ischnoplax Carpenter in Dall, 1879 x Ischnoplax pectinatus (Sowerby, 1832) C C C C C C C C x Género Stenoplax Carpenter in Dall, 1879 x Stenoplax bahamensis Kaas y Belle, 1987 R R x Stenoplax purpurascens (C. B. Adams, 1845) C C C C C C C C x Stenoplax boogii (Haddon, 1886) R R R R x Subfamilia CALLISTOPLACINAE Género Callistochiton Carpenter in Dall, 1879 x Callistochiton shuttleworthianus Pilsbry, 1893 C C C C C C C C x Género Ceratozona Dall, 1882 x Ceratozona squalida (C. -

Contributions in Science
LOS ANGELES CONTRIBUTIONS COUNTY MUSEUM IN SCIENCE NUMBER 189 MAY 4, 1970 REINSTATEMENT OF THE TURRID GENUS BELLASPIRA CONRAD, 1868 (MOLLUSCA: GASTROPODA) WITH A REVIEW OF THE KNOWN SPECIES By JAMES H. MCLEAN AND LEROY H. POORMAN Los ANGELES COUNTY MUSEUM OF NATURAL HISTORY • EXPOSITION PARK LOS ANGELES, CALIFORNIA 90007 CONTRIBUTIONS IN SCIENCE is a series of miscellaneous technical papers in the fields of Biology, Geology and Anthropology, published at irregular intervals by the Los Angeles County Museum of Natural History. Issues are numbered sep arately, and numbers run consecutively regardless of subject matter. Number 1 was issued January 23, 1957. The series is available to scientific institutions and scien tists on an exchange basis. Copies may also be purchased at a nominal price. Inquiries should be directed to Virginia D. Miller, Los Angeles County Museum of Natural History, 900 Exposition Boulevard, Los Angeles, California 90007. ROBERT J. LAVENBERG Managing Editor INSTRUCTIONS FOR AUTHORS Manuscripts for the LOS ANGELES COUNTY MUSEUM, CONTRIBU TIONS IN SCIENCE may be in any field of Life or Earth Sciences. Acceptance of papers will be determined by the amount and character of new information. Al though priority will be given to manuscripts by staff members, or to papers dealing largely with specimens in the Museum's collections, other technical papers will be considered. All manuscripts must be recommended for consideration by the curator in charge of the proper section or by the editorial board. Manuscripts must conform to those specifications listed below and will be examined for suitability by an Edi torial Committee. They may also be subject to review by competent specialists out side the Museum. -

Turridae (Mollusca: Gastropoda) of Southern Africa and Mozambique
Ann. Natal Mus. Vol. 29(1) Pages 167-320 Pietermaritzburg May, 1988 Turridae (Mollusca: Gastropoda) of southern Africa and Mozambique. Part 4. Subfamilies Drilliinae, Crassispirinae and Strictispirinae by R. N. Kilburn (Natal Museum, Pietermaritzburg) SYNOPSIS 71 species (42 previously undescribed) are covered: Drilliinae (33 species), Strictispirinae (3), Crassispirinae (35). New genera: Orrmaesia, Acinodrillia (Drilliinae); Inkinga (Strictispirinae); Naudedrillia, Nquma, Psittacodrillia, Calcatodrillia, Funa (Crassispirinae). New species: Drilliinae: Acinodrillia viscum, A. amazimba; Clavus groschi; Tylotiella isibopho, T. basipunctata, T. papi/io, T. herberti, T. sulekile, T. quadrata, Agladrillia ukuminxa, A. piscorum; Drillia (Drillia) spirostachys, D. (Clathrodrillia) connelli; Orrmaesia dorsicosta, O. nucella; Splendrillia mikrokamelos, S. kylix, S. alabastrum, S. skambos, S. sarda, S. daviesi. Crassispirinae: Crassiclava balteata; Inquisitor nodicostatus, /. arctatus, I. latiriformis, I. isabella, Funa fraterculus, F. asra; Naudedrillia filosa, N. perardua, N. cerea, N. angulata, N. nealyoungi, N. mitromorpha; Pseudexomilus fenestratus; Ceritoturris nataliae; Nquma scalpta; Haedropleura summa; Mauidrillia felina; Calcatodrillia chamaeleon, C. hololeukos; Buchema dichroma. New generic records: Clavus Montfort, 1810, Tylotiella Habe, 1958, Iredalea Oliver, 1915, Splendrillia Hedley, 1922, Agladrillia Woodring, 1928 (Drilliinae); Paradrillia Makiyama, 1940 (Strictispirinae); Crassiclava McLean, 1971, Pseudexomilus Powell, 1944, Haedropleura -

Interpreting the Paleozoogeography and Sea Level History of Thermally
Monographs of the Western North American Naturalist Volume 7 8th California Islands Symposium Article 6 9-9-2014 Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California Daniel R. Muhs U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Lindsey T. Groves Natural History Museum of Los Angeles County, [email protected] R. Randall Schumann U.S. Geological Survey, Denver Federal Center, Denver, CO, [email protected] Follow this and additional works at: https://scholarsarchive.byu.edu/mwnan Recommended Citation Muhs, Daniel R.; Groves, Lindsey T.; and Schumann, R. Randall (2014) "Interpreting the paleozoogeography and sea level history of thermally anomalous marine terrace faunas: a case study from the Last Interglacial Complex of San Clemente Island, California," Monographs of the Western North American Naturalist: Vol. 7 , Article 6. Available at: https://scholarsarchive.byu.edu/mwnan/vol7/iss1/6 This Monograph is brought to you for free and open access by the Western North American Naturalist Publications at BYU ScholarsArchive. It has been accepted for inclusion in Monographs of the Western North American Naturalist by an authorized editor of BYU ScholarsArchive. For more information, please contact [email protected], [email protected]. Monographs of the Western North American Naturalist 7, © 2014, pp. 82–108 INTERPRETING THE PALEOZOOGEOGRAPHY AND SEA LEVEL HISTORY OF THERMALLY ANOMALOUS MARINE TERRACE FAUNAS: A CASE STUDY FROM THE LAST INTERGLACIAL COMPLEX OF SAN CLEMENTE ISLAND, CALIFORNIA Daniel R. Muhs1,3, Lindsey T. Groves2, and R. Randall Schumann1 ABSTRACT.—Marine invertebrate faunas with mixtures of extralimital southern and extralimital northern faunal elements, called thermally anomalous faunas, have been recognized for more than a century in the Quaternary marine terrace record of the Pacific Coast of North America. -

Portadas 25 (1)
© Sociedad Española de Malacología Iberus, 28 (2): 1-4, 2010 Haedropleura ryalli, a new species from São Tomé Island (Gastropoda, Turridae) Haedropleura ryalli, nueva especie de la isla de São Tomé (Gastropoda, Turridae) Juan HORRO*, Sandro GORI** and Emilio ROLÁN*** Recibido el 10-III-2010. Aceptado el 31-VIII-2010 ABSTRACT A new species of the genus Haedropleura Bucquoy, Dautzenberg and Dollfus, 1883 from the island of São Tomé, West Africa, is described and figured. RESUMEN Se describe e ilustra una nueva especie del género Haedropleura Bucquoy, Dautzenberg y Dollfus, 1883 recolectada en la isla de São Tomé, África occidental. INTRODUCTION The genus Haedropleura Bucquoy, While studying the material collected Dautzenberg and Dollfus, 1883 is men- by the second author by scuba diving in tioned in POWELL (1966) who describes that island, the authors have identified a the shell, operculum and radula and new species of Haedropleura which is refers that this genus has species from described and illustrated in this paper. the European Miocene and Pliocene and also Recent from Europe and West Abbreviations: Africa down to South Africa. The introduction of scuba diving as MNHM Muséum national d’Histoire a sampling technique in recent times naturelle, Paris along several areas of the West African MNCN Museo Nacional de Ciencias coast where it had scarcely been used Naturales, Madrid before has led to the availability of MNHN Muséum national d’Histoire material from waters deeper than previ- naturelle, Paris ously studied ones, bringing a number ZMB Zoologisches Museum, Berlin of interesting new species to the atten- CJH collection Juan Horro, Vigo tion of malacologists. -
Catalog of Recent Molluscan Types in the Natural History Museum of Los
CATALOG OF RECENT MOLLUSCAN TYPES IN THE NATURAL HISTORY MUSEUM OF LOS ANGELES COUNTY compiled by Lindsey T. Groves NATURAL HISTORY MUSEUM OF LOS ANGELES COUNTY MALACOLOGY SECTION 900 Exposition Boulevard, Los Angeles, California, 90007, USA [email protected] [last updated 3 January, 2012] Abstract. Recent molluscan types currently housed in the Malacology Section of the Natural History Museum of Los Angeles County are listed herein. Each type lot entry includes original species/subspecies name, original genus, author, original citation, type status, LACM number, number of specimens and preservation, type locality or collection locality, collector [if known], date collected [if known], and current taxonomic status if different from original description. INTRODUCTION This catalog is published in accordance with Recommendation 72F.4 of the International Code of Zoological Nomenclature (Ride & others, 2000:79). This listing includes all molluscan type material currently housed at the Natural History Museum of Los Angeles County and supercedes the type listing of Sphon (1971). To date 1922 molluscan type lots (71,000+ specimens) are reported for 1147 species and 99 subspecies, which includes 589 holotypes, 1277 paratype lots, 10 lectotypes, 13 paralectotype lots, 29 syntype lots, 3 neotypes, and 2 neotype lots in 550+ references. Type categories listed include: Holotype (in red), paratype(s) (in blue), syntype(s) (in orange), lectotype (in pink), paralectotype(s) (in aqua), and neotype and/or neotype lots (in brown) as defined by Ride & others (2000). Because the term cotype is not recognized by the ICZN, lots originally designated as cotype(s) (in green) by original specimen labels are now referred to as syntype(s). -

Phylum Rhodophyta Class Florideophycyaea Order Corallinales Coralline Algae Gen
Upper Pliocene (Piacenzian) in the Reagin Collection Western Atlantic Eastern United States February 1, 2015 Legend Y - Zone 2 Yorktown Formation. North Carolina and Virginia D - Duplin Formation. North and South Carolina J - Jackson Bluff Formation. Florida Panhandle T - Tamiami Formation. South Florida C - Chowan River Formation. North Carolina R - Raysor Formation. South Carolina G - Goose Creek Limestone. South Carolina Phylum Rhodophyta Class Florideophycyaea Order Corallinales Coralline Algae gen. sp. indent. T Phylum Bryozoa Class Gymnolaemata Order Cheilostomata Tetrocycloecia sp. Y Family, Genus, species indet. YDJTC Family Hippoporinidae Hippoporidra edax (Busk, 1861) YDC Family Schizoporellidae Schizoporella sp. T Phylum Cnidaria Class Anthozoa Order Scleractinia Family Caryophyllidae Paracyathus vaughni Gane, 1985 T ` Family Faviidae Diploria clivosa (Ellis & Solander, 1786) T Diploria sarasotana Weisbord, 1974 Solenastrea bouroni Edwards & Haime, 1849 T Solenastrea hyades (Dana, 1846) Favia fragum? (Esper, 1868) T Family Oculinidae Oculina sarasotana Weisbord, 1974 T Family Rhizangiidae Septastrea bella (Conrad, 1841) C Septastrea marylandica (Conrad, 1837) YDJTC Septastrea crassa (Holmes, 1858) YT Astrangia leonensis Weisbord, 1974 T Family Siderastreidae Siderastrea dalli Vaughn, 1919 T Family Mussidae Syzgophyllia dentata (Duncan, 1863) T Family Pocilloporidae Stylophora granulata Duncan, 1863 T Phylum Brachiopoda Class Articulata Order Neotremata Family Discinidae Discinisca lugubris (Conrad, 1843) JT Phylum Mollusca Class -

Type Specimens in the Molluscan Collection of the Bailey-Matthews National Shell Museum, Florida, USA
Zootaxa 4951 (1): 001–040 ISSN 1175-5326 (print edition) https://www.mapress.com/j/zt/ Article ZOOTAXA Copyright © 2021 Magnolia Press ISSN 1175-5334 (online edition) https://doi.org/10.11646/zootaxa.4951.1.1 http://zoobank.org/urn:lsid:zoobank.org:pub:E2928477-FEC9-45CC-89C9-0B020D742EFA Type specimens in the molluscan collection of the Bailey-Matthews National Shell Museum, Florida, USA JOSÉ H. LEAL Bailey-Matthews National Shell Museum, 3075 Sanibel-Captiva Road, Sanibel, FL 33957, USA. �[email protected]; https://orcid.org/0000-0002-3786-0368 Abstract This article lists and comments on the primary and secondary types represented in the collection of the Bailey-Matthews National Shell Museum (BMSM), on Sanibel, Florida, USA. The collection includes 464 type specimens, of which 15 are holotypes, representing 149 taxa, of which 145 are species and four subspecies. The BMSM collection is fully catalogued and posted online via the Museum’s website, in addition to iDigBio and GBIF. The publication of this annotated list intends to improve on the accessibility and promote this important group of name-bearing specimens, which includes, among other cases, types originating from orphaned collections and material poorly documented in the original descriptions. Eighty-two types were selected for illustration, and the photos of all BMSM types are available as part of the BMSM online collection catalog. Key words: Nomenclature, taxonomy, biodiversity, malacology, holotype, paratype, Mollusca, Gastropoda, Bivalvia, Scaphopoda, types Introduction The Bailey-Matthews National Shell Museum opened to the public in July 1995, and was officially dedicated in No- vember 1995. Its first director, Dr.