Olivella Dama (Mawe)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Marine and Brackish Water Mollusca of the State of Mississippi
Gulf and Caribbean Research Volume 1 Issue 1 January 1961 The Marine and Brackish Water Mollusca of the State of Mississippi Donald R. Moore Gulf Coast Research Laboratory Follow this and additional works at: https://aquila.usm.edu/gcr Recommended Citation Moore, D. R. 1961. The Marine and Brackish Water Mollusca of the State of Mississippi. Gulf Research Reports 1 (1): 1-58. Retrieved from https://aquila.usm.edu/gcr/vol1/iss1/1 DOI: https://doi.org/10.18785/grr.0101.01 This Article is brought to you for free and open access by The Aquila Digital Community. It has been accepted for inclusion in Gulf and Caribbean Research by an authorized editor of The Aquila Digital Community. For more information, please contact [email protected]. Gulf Research Reports Volume 1, Number 1 Ocean Springs, Mississippi April, 1961 A JOURNAL DEVOTED PRIMARILY TO PUBLICATION OF THE DATA OF THE MARINE SCIENCES, CHIEFLY OF THE GULF OF MEXICO AND ADJACENT WATERS. GORDON GUNTER, Editor Published by the GULF COAST RESEARCH LABORATORY Ocean Springs, Mississippi SHAUGHNESSY PRINTING CO.. EILOXI, MISS. 0 U c x 41 f 4 21 3 a THE MARINE AND BRACKISH WATER MOLLUSCA of the STATE OF MISSISSIPPI Donald R. Moore GULF COAST RESEARCH LABORATORY and DEPARTMENT OF BIOLOGY, MISSISSIPPI SOUTHERN COLLEGE I -1- TABLE OF CONTENTS Introduction ............................................... Page 3 Historical Account ........................................ Page 3 Procedure of Work ....................................... Page 4 Description of the Mississippi Coast ....................... Page 5 The Physical Environment ................................ Page '7 List of Mississippi Marine and Brackish Water Mollusca . Page 11 Discussion of Species ...................................... Page 17 Supplementary Note ..................................... -

Las Familias Olividae Y Olivellidae De La Colección Malacológica De La Escuela Nacional De Ciencias Biológicas, IPN, México
Revista Mexicana de Biodiversidad 82: 1138-1144, 2011 Las familias Olividae y Olivellidae de la Colección Malacológica de la Escuela Nacional de Ciencias Biológicas, IPN, México The families Olividae and Olivellidae at the Malacological Collection of the Escuela Nacional de Ciencias Biológicas, IPN, Mexico Ricardo Pliego-Cárdenas1 y Aurora González-Pedraza2 1 Planta Experimental de Producción Acuícola, Universidad Autónoma Metropolitana Unidad Iztapalapa. Av. San Rafael Atlixco 186, Col. Vicentina, Apartado postal 55-535, México, D. F., México. 2 Laboratorio de Ecología, Escuela Nacional de Ciencias Biológicas, Instituto Politécnico Nacional. Prol. Carpio y Plan de Ayala s/n, Col. Casco de Santo Tomas 11340 México, D.F., México. [email protected] Resumen. Se presenta una evaluación de las conchas de los caracoles de las familias Olividae y Olivellidae de la colección malacológica de la Escuela Nacional de Ciencias Biológicas, IPN, México. Se revisaron 205 lotes y 1173 ejemplares. Olividae está representada por los géneros Oliva Bruguière, 1789 y Agaronia Gray, 1839 con 8 y 4 especies respectivamente, y Olivellidae por Olivella Swainson, 1831 y Jaspidella Olsson, 1956 con 15 y 2 especies, respectivamente. Se consideran nuevos registros de Agaronia griseoalba (von Martens, 1897) para el Pacífico mexicano. Un registro de A. nica López, Montoya y López, 1988 sugiere una ampliación de su distribución geográfica. Palabras clave: neogastrópodos, nuevo registro, ámbito de distribución. Abstract. We present an evaluation of the shells from sea snails of the families Olividae and Olivellidae deposited at the mollusk collection of the Escuela Nacional de Ciencias Biológicas, IPN, México. Two hundred and five lots and 1173 specimens were analyzed. -
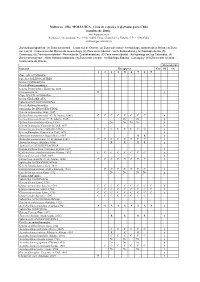
Moluscos - Filo MOLLUSCA
Moluscos - Filo MOLLUSCA. Lista de especies registradas para Cuba (octubre de 2006). José Espinosa Sáez Instituto de Oceanología, Ave 1ª No. 18406, Playa, Ciudad de La Habana, C.P. 11200, Cuba [email protected] Zonas biogeográficas: (1) Zona suroriental – Costa sur de Oriente, (2) Zona surcentral - Archipiélago Jardines de la Reina, (3) Zona sur central - Costa al sur del Macizo de Guamuhaya, (4) Zona suroccidental - Golfo de Batabanó y Archipiélago de los, (5) Canarreos, (6) Zona suroccidental - Península de Guanahacabibes, (7) Zona noroccidental - Archipiélago de Los Colorados, (8) Zona noroccidental - Norte Habana-Matanzas, (9) Zona norte-central - Archipiélago Sabana - Camagüey, (10) Zona norte-oriental - Costa norte de Oriente Abreviaturas Especies Bioegiones Cu Pl Oc 1 2 3 4 5 6 7 8 9 Clase APLACOPHORA Subclase SOLENOGASTRES Orden CAVIBELONIA Familia Proneomeniidae Género Proneomenia Hubrecht, 1880 Proneomenia sp . R x Clase POLYPLACOPHORA Orden NEOLORICATA Suborden ISCHNOCHITONINA Familia Ischnochitonidae Subfamilia ISCHNOCHITONINAE Género Ischnochiton Gray, 1847 Ischnochiton erythronotus (C. B. Adams, 1845) C C C C C C C C x Ischnochiton papillosus (C. B. Adams, 1845) Nc Nc x Ischnochiton striolatus (Gray, 1828) Nc Nc Nc Nc x Género Ischnoplax Carpenter in Dall, 1879 x Ischnoplax pectinatus (Sowerby, 1832) C C C C C C C C x Género Stenoplax Carpenter in Dall, 1879 x Stenoplax bahamensis Kaas y Belle, 1987 R R x Stenoplax purpurascens (C. B. Adams, 1845) C C C C C C C C x Stenoplax boogii (Haddon, 1886) R R R R x Subfamilia CALLISTOPLACINAE Género Callistochiton Carpenter in Dall, 1879 x Callistochiton shuttleworthianus Pilsbry, 1893 C C C C C C C C x Género Ceratozona Dall, 1882 x Ceratozona squalida (C. -

Current Status of Philippine Mollusk Museum Collections and Research, and Their Implications on Biodiversity Science and Conservation
Philippine Journal of Science 147 (1): 123-163, March 2018 ISSN 0031 - 7683 Date Received: 28 Feb 2017 Current Status of Philippine Mollusk Museum Collections and Research, and their Implications on Biodiversity Science and Conservation Dino Angelo E. Ramos2*, Gizelle A. Batomalaque1,3, and Jonathan A. Anticamara1,2 1Ecology and Taxonomy Academic Group (ETAG), Institute of Biology, University of the Philippines, Diliman, Quezon City 1101 Philippines 2UP Biology Invertebrate Museum, Institute of Biology, University of the Philippines, Diliman, Quezon City 1101 Philippines 3Department of Biodiversity, Earth, and Environmental Science, Drexel University, Philadelphia, Pennsylvania 19104 USA Mollusks are an invaluable resource in the Philippines, but recent reviews on the status of museum collections of mollusks or research trends in the country are lacking. Such assessments can contribute to a more comprehensive evaluation of natural history museums in the Philippines, as well as biodiversity management. This review showed that local museums in the Philippines have much to improve in terms of their accessibility and geographic coverage in order to effectively cater to research and conservation needs of the country. Online access to databases was lacking for local museums, making it cumbersome to retrieve collection information. The UST museum held the most species and subspecies across all museums (4899), comparable to the national museums of countries such as the USA and France. In terms of size, there were larger Philippine mollusk collections in museums abroad. Majority of mollusk specimens come from Regions 4 and 7, while the CAR and Region 12 were least sampled. Publications on Philippine mollusks are dominated by taxonomic and biodiversity research. -

Sociedad Malacológica De Chile
AMICI MOLLUSCARUM Número 20(1), año 2012 Sociedad Malacológica de Chile AMICI MOLLUSCARUM Número 20(1)20(1),, año 2012012222 Amici Molluscarum es una revista de publicación anual bilingüe, editada por la Sociedad Malacológica de Chile (SMACH) desde el año 1992, siendo la continuación del boletín Comunicaciones , publicado entre 1979 y 1986. Cuenta con el patrocinio del Museo Nacional de Historia Natural de Chile (MNHNCL). Tiene el propósito de publicar artículos científicos originales, así como también comunicaciones breves (notas científicas), fichas de especies, comentarios de libros y revisiones en todos los ámbitos de la malacología. ISSN 07180718----97619761 (versión en línea) Los textos e ilustraciones contenidos en esta revista pueden reproducirse, siempre que se mencione su origen, indicando el nombre del autor o su procedencia, y se agregue el volumen y año de publicación. Imagen de la cubierta: Vista interna y externa de valva derecha de un ejemplar de Diplodon chilensis (D. Jackson y D. Jackson) Imagen de la contracubierta: Vista ventral y dorsal de Aylacostoma chloroticum (R.E. Vogler). Amici Molluscarum http://www.amicimolluscarum.com Sociedad Malacológica de Chile (SMACH) http://www.smach.cl AMICI MOLLUSCARUM Sociedad Malacológica de Chile (SMACH) Comité editorial Editor jefe Gonzalo Collado Universidad de Chile, Santiago, Chile Editor de producción Cristian Aldea Fundación CEQUA, Punta Arenas, Chile Editores asociados Omar Ávila-Poveda Universidad del Mar, Oaxaca, México Roberto Cipriani California State University, Fullerton, -

Trade in Molluskan Religiofauna Between the Southwe Stern
TRADE IN MOLLUSKAN RELIGIOFAUNA BETWEEN THE SOUTHWE STERN UNITED STATES AND SOUTHERN CALIFORNIA by WILLIAM HOYT SMITH A DISSERTATION Pres ented to the Department of Anthropology and the Graduate School of the University of Oregon in partial fulfillment of the requirements for the degree of Doctor of Philosophy June 2002 --------------- ---- ii "Tr ade in Molluskan Religiofauna Between the Southwestern United States and Southern California ," a dissertation prepared by William Hoyt Smith in partial fulfillment of the requirements for the Doctor of Philosophy degree in the Department of Anthropology. This dissertation has been approved and accepted by: Dr . C. a ining Committee Committee in Charge : Dr. C. Melvin Aikens Dr. Jon Erlandson Dr. Don Dumond Dr. Leland Roth Dean of the Graduate School L iv An Abstract of the Dissertation of William Smith for the Degree of Doctor of Philosophy the Department Anthropology to be taken June 2002 Title: TRADE IN MOLLUSKAN RELIGIOFAUNA BETWEEN THE SOUTHWESTERN UNITED STATES AND SOUTHERN CALIFORNIA Approved: Dr. C. Melvin Aikens This dissertation describes prehistoric trade between Native American groups of the North American Southwest and southern California. Archaeologic and ethnographic sources have been used to create a major new quant ive database on Southwest-southern California trade. I draw on this data base to characterize the routes and intensity of exchange, as well as speci objects involved in native trade networks. facts made of Olivella and Halio s shells were traded eastward from the southern California Channel Islands into the Anasazi, Mogollon, and Hohokam regions of the Southwest in south Nevada, Arizona, and New Mexico. -

Diversity and Community Composition of Marine Mollusks Fauna on a Mainland Island of the Coast of Paraná, Southern Brazil
Pesquisa e Ensino em Ciências ISSN 2526-8236 (online edition) Exatas e da Natureza Pesquisa e Ensino em Ciências 2(1): 48–59 (2018) ARTICLE Research and Teaching in Exatas e da Natureza © 2018 UFCG / CFP / UACEN Exact and Natural Sciences Diversity and community composition of marine mollusks fauna on a mainland island of the coast of Paraná, southern Brazil Marcos de Vasconcellos Gernet1, Eduardo Colley2, Elizângela da Veiga Santos1,3 & Carlos João Birckolz1 (1) Universidade Federal do Paraná, Laboratório de Ecologia Aplicada e Bioinvasões, Rua Rio Grande do Norte 145, Mirassol 83255-000, Pontal do Paraná, Paraná, Brazil. E-mail: [email protected], [email protected] (2) Universidade de São Paulo, Museu de Zoologia, Avenida Nazaré 481, Ipiranga 04263-000, São Paulo, São Paulo, Brazil. E-mail: [email protected] (3) Universidade Federal do Paraná, Setor Litoral, Rua Jaguariaíva, 512, Caiobá 83260-000, Matinhos, Paraná, Brazil. E-mail: [email protected] Gernet M.V., Colley E., Santos E.V. & Birckolz C.J. (2018) Diversity and community composition of marine mollusks fauna on a mainland island of the coast of Paraná, southern Brazil. Pesquisa e Ensino em Ciências Exatas e da Natureza, 2(1): 48–59. http://dx.doi.org/10.29215/pecen.v2i1.580 Diversidade e composição da comunidade de moluscos marinhos de uma ilha continental do litoral do Paraná, sul do Brasil Resumo: A Ilha do Farol é uma ilha continental, no estado do Paraná, sul do Brasil. Devido à sua posição em relação ao continente, três ambientes distintos são observados nela: área de costão rochoso exposto ao mar aberto (A); área estuarina (B); área de praia arenosa (C). -

A Taxonomic Review of California Holocene Callianax (Olivellidae: Gastropoda: Mollusca) Based on Shell Characters
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/339955665 A taxonomic review of California Holocene Callianax (Olivellidae: Gastropoda: Mollusca) based on shell characters Article · March 2020 CITATIONS READS 0 9 3 authors, including: Charles L. Powell David Berschauer Emeritus United States Geological Survey University of California, Irvine 100 PUBLICATIONS 339 CITATIONS 31 PUBLICATIONS 33 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Biodiversity Studies View project Pleistocene Marine Mollusks of California and Baja California: An Annotated Bibliography, 1866-2000, version 1 View project All content following this page was uploaded by Charles L. Powell on 16 March 2020. The user has requested enhancement of the downloaded file. A taxonomic review of California Holocene Callianax (Olivellidae. Gastropoda. Mollusca) based on shell characters Charles L. Powell II, Fred Vervaet and David Berschauer Supplemental Publication of the San Diego Shell Club ISSN 0738-9388 T H E F E S T I V U S 1 A publication of the San Diego Shell Club SUPPLEMENT March 2020 SPECIAL ISUE CLUB OFFICERS MISSION STATEMENT President Roger N. Clark The San Diego Shell Club was founded in 1961 as a non- Larry Buck Larry Buck Vice-President Bill Schramm profit organization for educational and scientific purposes. Corresp. Secretary Dave Michaelski More particularly to enjoy, study and promote the Recording Secretary Leo Kempczenski conservation of Mollusca and associated marine life Treasurer “Duffy” Daughenbaugh through lectures, club meetings and field trips. Our Past President Lisa Dawn Lindahl membership is diverse and includes beginning collectors, scientists, divers, underwater photographers and dealers. -

Diversidad De La Comunidad De Moluscos De Fondos Blandos Del Archipiélago Espíritu Santo, Golfo De California, México
INSTITUTO POLITÉCNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS DIVERSIDAD DE LA COMUNIDAD DE MOLUSCOS DE FONDOS BLANDOS DEL ARCHIPIÉLAGO ESPÍRITU SANTO, GOLFO DE CALIFORNIA, MÉXICO. TESIS QUE PARA OBTENER EL GRADO DE MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS PRESENTA ALEJANDRO BOSCH CALLAR LA PAZ, B.C.S., JUNIO DEL 2018 AGRADECIMIENTOS Primero que todo a mis padres por confiar en mí y apoyarme en todo momento de mi carrera. A mi esposa Susana Perera Valderrama por estar a mi lado en todo momento y por darme el mejor regalo que se puede esperar en la vida: mi hijo Alejandro Jr. Bosch Perera A mi hermana que a pesar de la distancia sigue estando presente para mi y sabe que la quiero con la vida. Al Centro Interdisciplinario de Ciencias Marinas por permitirme la realización de este trabajo en sus instalaciones. A mis hermano Franklin con quien emprendí este largo viaje fuera de nuestra tierra, por los tantos momentos, buenos y malos, y no cambiar a pesar de las dificultades. A mis hermanos los simbiontes que una ves mas nos reunimos pese a la distancia que nos separaba y nuestra amistad se fortalece cada día mas. Un agradecimiento especial a Angel de León Espinosa, por ser la primera persona en brindarme su apoyo cuando recién llegue a México, y por las muchas aventuras que nos hemos aventado y las que vendrán. Un Agradecimiento especial a Ramón y a Tatiana de Dream Yatch Charter por su apoyo en todo este tiempo que ha durado mi estancia aquí en La Paz. -

Inventory, Monitoring and Impact Assessment of Marine Biodiversity in the Seri Indian Territory, Gulf of California, Mexico
INVENTORY, MONITORING AND IMPACT ASSESSMENT OF MARINE BIODIVERSITY IN THE SERI INDIAN TERRITORY, GULF OF CALIFORNIA, MEXICO by Jorge Torre Cosío ________________________ A Dissertation Submitted to the Faculty of the SCHOOL OF RENEWABLE NATURAL RESOURCES In Partial Fulfillment of the Requirements For the Degree of DOCTOR OF PHYLOSOPHY WITH A MAJOR IN RENEWABLE NATURAL RESOURCES STUDIES In the Graduate College THE UNIVERSITY OF ARIZONA 2 0 0 2 Sign defense sheet STATEMENT BY AUTHOR This dissertation has been submitted in partial fulfillment of requirements for an advanced degree at The University of Arizona and is deposited in the University Library to be made available to borrowers under rules of the Library. Brief quotations from this dissertation are allowable without special permission, provided that accurate acknowledgment of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in his or her judgment the proposed use of the material is in the interests of scholarship. In all other instances, however, permission must be obtained from the author. SIGNED:____________________________ ACKNOWLEDGMENTS The Consejo Nacional de Ciencia y Tecnología (CONACyT) and the Wallace Research Foundation provided fellowships to the author. The Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) (grant FB463/L179/97), World Wildlife Fund (WWF) México and Gulf of California Program (grants PM93 and QP68), and the David and Lucile Packard Foundation (grant 2000-0351) funded this study. Administrative support for part of the funds was through Conservation International – México (CIMEX) program Gulf of California Bioregion and Conservación del Territorio Insular A.C. -

Checklist of Species Within the CCBNEP Study Area: References, Habitats, Distribution, and Abundance
Current Status and Historical Trends of the Estuarine Living Resources within the Corpus Christi Bay National Estuary Program Study Area Volume 4 of 4 Checklist of Species Within the CCBNEP Study Area: References, Habitats, Distribution, and Abundance Corpus Christi Bay National Estuary Program CCBNEP-06D • January 1996 This project has been funded in part by the United States Environmental Protection Agency under assistance agreement #CE-9963-01-2 to the Texas Natural Resource Conservation Commission. The contents of this document do not necessarily represent the views of the United States Environmental Protection Agency or the Texas Natural Resource Conservation Commission, nor do the contents of this document necessarily constitute the views or policy of the Corpus Christi Bay National Estuary Program Management Conference or its members. The information presented is intended to provide background information, including the professional opinion of the authors, for the Management Conference deliberations while drafting official policy in the Comprehensive Conservation and Management Plan (CCMP). The mention of trade names or commercial products does not in any way constitute an endorsement or recommendation for use. Volume 4 Checklist of Species within Corpus Christi Bay National Estuary Program Study Area: References, Habitats, Distribution, and Abundance John W. Tunnell, Jr. and Sandra A. Alvarado, Editors Center for Coastal Studies Texas A&M University - Corpus Christi 6300 Ocean Dr. Corpus Christi, Texas 78412 Current Status and Historical Trends of Estuarine Living Resources of the Corpus Christi Bay National Estuary Program Study Area January 1996 Policy Committee Commissioner John Baker Ms. Jane Saginaw Policy Committee Chair Policy Committee Vice-Chair Texas Natural Resource Regional Administrator, EPA Region 6 Conservation Commission Mr. -

Faunal Remains from Archaeology Sites in Southwestern New Mexico
University of New Mexico UNM Digital Repository Occasional Papers Museum of Southwestern Biology 9-21-2020 Faunal remains from archaeology sites in southwestern New Mexico Karen Gust Schollmeyer Archaeology Southwest S. O. MacDonald University of New Mexico Follow this and additional works at: https://digitalrepository.unm.edu/occasionalpapers Part of the Biodiversity Commons, Ecology and Evolutionary Biology Commons, and the Paleobiology Commons Recommended Citation PUBLICATIONS OF THE MUSEUM OF SOUTHWESTERN BIOLOGY UNIVERSITY OF NEW MEXICO https://digitalrepository.unm.edu/msb/ The publications of the Museum of Southwestern Biology of the University of New Mexico are issued under the auspices of the Dean of the College of Arts and Sciences and the Director of the Museum of Southwestern Biology. Short research studies that are specimen based are published as Occasional Papers, whereas longer contributions appear as Special Publications. All are peer-reviewed, numbered separately, and published on an irregular basis. The preferred format for citing the Museum's publications is Occasional Paper (or Special Publication), Museum of Southwestern Biology, The University of New Mexico, Albuquerque, NM. Authors interested in submitting scholarly work for publication should first contact the Managing ditE or, Museum Publications, Museum of Southwestern Biology for instructions. ©2020. The Museum of Southwestern Biology, The University of New Mexico, Albuquerque, NM 87131-0001, USA. ISSN 2380-4580 Managing Editor, Joseph Cook Subject Editor, Emily Jones This Article is brought to you for free and open access by the Museum of Southwestern Biology at UNM Digital Repository. It has been accepted for inclusion in Occasional Papers by an authorized administrator of UNM Digital Repository.