EEG and Sleep Effects of Tramadol Suggest Potential Antidepressant Effects with Different Mechanisms of Action
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Music Genre Preference and Tempo Alter Alpha and Beta Waves in Human Non-Musicians
Page 1 of 11 Impulse: The Premier Undergraduate Neuroscience Journal 2013 Music genre preference and tempo alter alpha and beta waves in human non-musicians. Nicole Hurless1, Aldijana Mekic1, Sebastian Peña1, Ethan Humphries1, Hunter Gentry1, 1 David F. Nichols 1Roanoke College, Salem, Virginia 24153 This study examined the effects of music genre and tempo on brain activation patterns in 10 non- musicians. Two genres (rock and jazz) and three tempos (slowed, medium/normal, and quickened) were examined using EEG recording and analyzed through Fast Fourier Transform (FFT) analysis. When participants listened to their preferred genre, an increase in alpha wave amplitude was observed. Alpha waves were not significantly affected by tempo. Beta wave amplitude increased significantly as the tempo increased. Genre had no effect on beta waves. The findings of this study indicate that genre preference and artificially modified tempo do affect alpha and beta wave activation in non-musicians listening to preselected songs. Abbreviations: BPM – beats per minute; EEG – electroencephalography; FFT – Fast Fourier Transform; ERP – event related potential; N2 – negative peak 200 milliseconds after stimulus; P3 – positive peak 300 milliseconds after stimulus Keywords: brain waves; EEG; FFT. Introduction For many people across cultures, music The behavioral relationship between is a common form of entertainment. Dillman- music preference and other personal Carpentier and Potter (2007) suggested that characteristics, such as those studied by music is an integral form of human Rentfrow and Gosling (2003), is evident. communication used to relay emotion, group However, the neurological bases of preference identity, and even political information. need to be studied more extensively in order to Although the scientific study of music has be understood. -

Quantitative EEG (QEEG) Analysis of Emotional Interaction Between Abusers and Victims in Intimate Partner Violence: a Pilot Study
brain sciences Article Quantitative EEG (QEEG) Analysis of Emotional Interaction between Abusers and Victims in Intimate Partner Violence: A Pilot Study Hee-Wook Weon 1, Youn-Eon Byun 2 and Hyun-Ja Lim 3,* 1 Department of Brain & Cognitive Science, Seoul University of Buddhism, Seoul 08559, Korea; [email protected] 2 Department of Youth Science, Kyonggi University, Suwon 16227, Korea; [email protected] 3 Department of Community Health & Epidemiology, University of Saskatchewan, Saskatoon, SK S7N 2Z4, Canada * Correspondence: [email protected] Abstract: Background: The perpetrators of intimate partner violence (IPV) and their victims have different emotional states. Abusers typically have problems associated with low self-esteem, low self-awareness, violence, anger, and communication, whereas victims experience mental distress and physical pain. The emotions surrounding IPV for both abuser and victim are key influences on their behavior and their relationship. Methods: The objective of this pilot study was to examine emotional and psychological interactions between IPV abusers and victims using quantified electroencephalo- gram (QEEG). Two abuser–victim case couples and one non-abusive control couple were recruited from the Mental Image Recovery Program for domestic violence victims in Seoul, South Korea, from Citation: Weon, H.-W.; Byun, Y.-E.; 7–30 June 2017. Data collection and analysis were conducted using BrainMaster and NeuroGuide. Lim, H.-J. Quantitative EEG (QEEG) The emotional pattern characteristics between abuser and victim were examined and compared to Analysis of Emotional Interaction those of the non-abusive couple. Results: Emotional states and reaction patterns were different for between Abusers and Victims in the non-abusive and IPV couples. -

Tramadol (Ultram)
TRAMADOL (ULTRAM) Tramadol is FDA approved for the treatment of musculoskeletal pain. Studies have shown it is useful in treating the pain associated with diabetic neuropathy and other pain conditions. Tramadol comes in 50 mg tablets. The maximum dose is two tablets four times per day unless your kidney function is below normal or you are over 75 years old, in which case the maximum dose is two tablets three times per day. The main side effects of Tramadol are drowsiness, sedation, and stomach upset, all of which are minimized by slowly raising the dose. About 5% of patients have stomach upset at any dose of Tramadol and cannot take the medicine. Other risks include seizures (occur in less than 1/100,000 and are more likely if you have seizures) and possibly abuse (relevant if you have abused drugs in the past). Tramadol should be started at a low dose and raise the dose slowly toward the maximum dose. Start with one tablet at bedtime. After 3 - 7 days, increase to one tablet twice daily (morning and bedtime). After an additional 3 - 7 days, increase to one tablet three times per day (morning, noon, and bedtime). After an additional 3 - 7 days, increase to one tablet four times per day (1 tablet with each meal and 1 at bedtime). At that point, the dose may be increased or adjusted depending on how you are doing. To increase further, you will: Add a second tablet at bedtime (one tablet three times per day and two tablets at bedtime). After 3 - 7 days, add a second tablet to another dose (one tablet twice per day and two tablets twice per day). -

Effects of Prophylactic Ketamine and Pethidine to Control Postanesthetic Shivering: a Comparative Study
Biomedical Research and Therapy, 5(12):2898-2903 Original Research Effects of prophylactic ketamine and pethidine to control postanesthetic shivering: A comparative study Masoum Khoshfetrat1, Ali Rosom Jalali2, Gholamreza Komeili3, Aliakbar Keykha4;∗ ABSTRACT Background: Shivering is an undesirable complication following general anesthesia and spinal anesthesia, whose early control can reduce postoperative metabolic and respiratory complications. Therefore, this study aims to compare the effects of prophylactic injection of ketamine and pethi- dine on postoperative shivering.Methods: This double-blind clinical trial was performed on 105 patients with short-term orthopedic and ENT surgery. The patients were randomly divided into three groups; 20 minutes before the end of the surgery, 0.4 mg/kg of pethidine was injected to the first group, 0.5 mg/kg of ketamine was injected to the second group, and normal saline was injected to the third group. After the surgery, the tympanic membrane temperature was measured at 0, 10, 20, and 30 minutes. The shivering was also measured by a four-point grading from zero (no shiv- ering) to four (severe shivering). Data were analyzed by one-way ANOVA, Kruskal Wallis, Chi-square 1Doctor of Medicine (MD), Fellow of and Pearson correlation. Results: The mean age of patients was 35.811.45 years in the ketamine Critical Care Medicine (FCCM), group, 34.811.64 years in the normal saline group, and 33.1110.5 years in the pethidine group. Department of Anesthesiology and The one-way ANOVA showed no significant difference in the mean age between the three groups Critical Care, Khatam-Al-Anbiya (P=0.645). -

Current Awareness in Clinical Toxicology Editors: Damian Ballam Msc and Allister Vale MD
Current Awareness in Clinical Toxicology Editors: Damian Ballam MSc and Allister Vale MD February 2016 CONTENTS General Toxicology 9 Metals 38 Management 21 Pesticides 41 Drugs 23 Chemical Warfare 42 Chemical Incidents & 32 Plants 43 Pollution Chemicals 33 Animals 43 CURRENT AWARENESS PAPERS OF THE MONTH How toxic is ibogaine? Litjens RPW, Brunt TM. Clin Toxicol 2016; online early: doi: 10.3109/15563650.2016.1138226: Context Ibogaine is a psychoactive indole alkaloid found in the African rainforest shrub Tabernanthe Iboga. It is unlicensed but used in the treatment of drug and alcohol addiction. However, reports of ibogaine's toxicity are cause for concern. Objectives To review ibogaine's pharmacokinetics and pharmacodynamics, mechanisms of action and reported toxicity. Methods A search of the literature available on PubMed was done, using the keywords "ibogaine" and "noribogaine". The search criteria were "mechanism of action", "pharmacokinetics", "pharmacodynamics", "neurotransmitters", "toxicology", "toxicity", "cardiac", "neurotoxic", "human data", "animal data", "addiction", "anti-addictive", "withdrawal", "death" and "fatalities". The searches identified 382 unique references, of which 156 involved human data. Further research revealed 14 detailed toxicological case reports. Current Awareness in Clinical Toxicology is produced monthly for the American Academy of Clinical Toxicology by the Birmingham Unit of the UK National Poisons Information Service, with contributions from the Cardiff, Edinburgh, and Newcastle Units. The NPIS is commissioned by Public Health England Current Awareness in Clinical Toxicology Editors: Damian Ballam MSc and Allister Vale MD February 2016 Current Awareness in Clinical Toxicology is produced monthly for the American Academy of Clinical Toxicology by the Birmingham Unit of the UK National Poisons Information Service, with contributions from the Cardiff, Edinburgh, and Newcastle Units. -

Quantitative Drug Test Menu Section 2
1 Guthrie Square, Sayre, PA 18840 Bill To: Client GMG Toxicology Laboratory Requisition Toll Free Phone (844) 617-4719 Insurance Request Date: _____/______/______ Medical Director: Hani Hojjati, MD Fax (570) 887-4729 Patient PATIENT INFORMATION (PLEASE PRINT IN BLACK INK) INSURANCE BILLING INFORMATION (PLEASE PRINT IN BLACK INK) Pt Last Name First M I PRIMARY Medicare Medicaid Other Ins. Self Spouse Child __ Subscriber Last Name First M Address Birth Date Sex M F Beneficiary/Member # Group # City Pt. SS# or MRN Claims Name and Address City ST ZIP ST ZIP Home Phone (Attach a copy of the patient's insurance card and information) SECONDARY Medicare Medicaid Other Ins. Self Spouse Child Employer Work Phone Subscriber Last Name First M Work Address City ST ZIP Beneficiary/Member # Group # __ CLIENT INFORMATION - REFERRING PHYSICIAN Claims Name and Address City ST ZIP Client Address: (Atttach a copy of the patient's insurance card and information) COLLECTION / REPORTING INFORMATION Copy to: FAX Results to __ CALL Results to Phone: Fax: Date Collected: Time Collected: AM PM Specimen Type: Urine Saliva Other ___________________ Physician Signature (legible - No Stamp) For Lab Use Only (Required for Medicare & Medicaid patient orders) Signed ABN Obtained Place Lab Label Here Contact Laboratory Medical Director (570-887-4719) with questions concerning medical necessity PHYSICIAN When ordering tests, the physician is required to make an independent medical necessity decision with regard to each test thelaboratory will bill. The physician also understands he or she is required NOTICE to (1) submit ICD-10 diagnosis supported in the patient's medical record as documentation of the medical necessity or (2) explain and have the patient sign an ABN. -

Procedure Manual for Polysomnography
Procedure Manual for Polysomnography The PSG Reading Center Case Western Reserve University Triangle Building 11400 Euclid Avenue Suite 260 Cleveland, Ohio 44106 January 7-9, 2002 (Rev. 8/20/02) Table of Contents 1.0 INTRODUCTION 1.1 Definition of Sleep Apnea 1.2 Polysomnography 1.2.1 Signal Types 1.2.2 Sleep Stages 1.2.3 Respiratory Monitoring - Measurement Tools 1.3 Home Polysomnography - Sleep System 1.4 Glossary of Sleep Terms 2.0 HOME POLYSOMNOGRAPHY (PSG) 2.1 Supply List 2.1.1 Understanding the Electrode 2.1.1.1 Gold disks - Cleaning, Disinfecting, Conditioning 2.1.2 Cleaning and Disinfecting Other Sensors and Equipment 2.2 Preparation Pre-Visit Hook-up 2.3 Detailed Hookup Procedures 2.3.1 Setting Up in the Home 2.3.2 Sensor Placement Step 1: ECG Electrodes Step 2: Respiratory Bands Step 3: EEG Scalp Electrodes Preparation of Electrode Sites Attaching Gold Electrodes Step 4: Position Sensor Step 5: Oximier Step 6: Nasal Cannula Step 7: Thermistor Step 8 Leg Sensors 2.4 Checking Impedances and Signal Quality 2.4.1 Verify Connections and Auto Start On 2.4.2 Impedance Checks (Signal Verification Form - SV) 2.4.3 View Signals 2.5 Final Instructions to Participant and Morning After Procedures 2.6 Troubleshooting Equipment and Signal Quality 3.0 PSG DATA COLLECTION PROCEDURES 3.1 Compumedics Programs Used for Data Collection 3.1.1 Data Card Manager (Setting up Flashcard) 3.1.2 Net Beacon (PSG on Line) 3.1.3 Profusion Study Manager 3.1.4 Profusion PSG 3.2 PSG Sleep Data Retrieval Procedures 3.3 Backup Studies to Zip Cartridges 3.4 Review -

CAN YOU TAKE TRAMADOL with NEFOPAM Can You Take Tramadol with Nefopam
CAN YOU TAKE TRAMADOL WITH NEFOPAM can you take tramadol with nefopam tramadol 37 5 vs percocet 5 325 ultram tramadol pictures tramadol hcl tabs 50 mg tramadol 200 mg recreational drugs and heart can tramadol and percocet be mixed hbs robaxin tramadol interaction generic tramadol 319 immediate release how long tramadol stay in your urine does tramadol make you sleepy or awake tramadol acetaminophen\/codeine 120 12mg sol b tracert ex tramadol dosage for adults meloxicam/tramadol/amitriptyline/lidocaine/prilocaine apo tramadol high feeling on hydrocodone tramadol apteka internetowa olmed order tramadol/paracetamol from mexico tramadol quizlet flashcards microbiology tramadol has mu opioid agonist activity director jobs tramadol met ritalin sr strengths hur ta tramadol withdrawal in dogs tramadol te gebruiken bij tramadol dosis cachorros bulldog 2015 100mg tramadol 10mg hydrocodone images 100 tramadol termasuk jenis obat apa acyclovir side how to get rid of a tramadol high 200 ml tramadol withdrawal timeline drug interactions between percocet and tramadol comparison tramadol e morfina presentacion de tres can tramadol be taken with paracetamol indication and action tramadol review article template with photos tramadol codeine allergy rash best price tramadol online tramadol 93 58 dosage for ibuprofen tramadol v oxycodone pill colors can tramadol make you drowsy doll b tracert ex tramadol addiction withdrawal tramadol instant release oxycontin pictures can you drink wine with tramadol i can function tramadol hydrochloride sleepy tramadol cva -

Parkinson's Disease Fact Sheet
Parkinson’s Disease Fact Sheet About Parkinson’s Disease Parkinson’s disease is a progressive, incurable neurological disorder associated with a loss of dopamine-generating cells in the brain. It is primarily associated with progressive loss of motor control, but it results in a complex array of symptoms, including many non-motor symptoms. Parkinson’s impacts an estimated one million people in the United States. Critical Clinical Care Considerations • To avoid serious side effects, Parkinson’s patients need their medications on time, every time — do not skip or postpone doses. • Write down the exact times of day medications are to be administered so that doses are given on the same schedule the patient follows at home. • Do not substitute Parkinson’s medications or stop levodopa therapy abruptly. • Resume medications immediately following procedures, unless vomiting or severely incapacitated. • If an antipsychotic is necessary, use pimavanserin (Nuplazid), quetiapine (Seroquel) or clozapine (Clozaril). • Be alert for symptoms of dysphagia (trouble swallowing) and risk of pneumonia. • Ambulate as soon as medically safe. Patients may require assistance. Common Symptoms of Parkinson’s Disease Motor Non-Motor • Shaking or tremor at rest • Depression • Bradykinesia or freezing (being stuck • Anxiety in place when attempting to walk) • Constipation • Low voice volume or muffled speech • Cognitive decline and dementia • Lack of facial expression • Impulse control disorders • Stiffness or rigidity of the arms, legs • Orthostatic hypotension or -

Michelia Essential Oil Inhalation Increases Fast Alpha Wave Activity
Scientia Pharmaceutica Article Michelia Essential Oil Inhalation Increases Fast Alpha Wave Activity Phanit Koomhin 1,2,3,*, Apsorn Sattayakhom 2,4, Supaya Chandharakool 4, Jennarong Sinlapasorn 4, Sarunnat Suanjan 4, Sarawoot Palipoch 1, Prasit Na-ek 1, Chuchard Punsawad 1 and Narumol Matan 2,5 1 School of Medicine, Walailak University, Nakhonsithammarat 80160, Thailand; [email protected] (S.P.); [email protected] (P.N.-e.); [email protected] (C.P.) 2 Center of Excellence in Innovation on Essential oil, Walailak University, Nakhonsithammarat 80160, Thailand; [email protected] (A.S.); [email protected] (N.M.) 3 Research Group in Applied, Computational and Theoretical Science (ACTS), Walailak University, Nakhonsithammarat 80160, Thailand 4 School of Allied Health Sciences, Walailak University, Nakhonsithammarat 80160, Thailand; [email protected] (S.C.); [email protected] (J.S.); [email protected] (S.S.) 5 School of Agricultural Technology, Walailak University, Nakhonsithammarat 80160, Thailand * Correspondence: [email protected]; Tel.: +66-95295-0550 Received: 13 February 2020; Accepted: 6 May 2020; Published: 9 May 2020 Abstract: Essential oils are volatile fragrance liquids extracted from plants, and their compound annual growth rate is expected to expand to 8.6% from 2019 to 2025, according to Grand View Research. Essential oils have several domains of application, such as in the food and beverage industry, in cosmetics, as well as for medicinal use. In this study, Michelia alba essential oil was extracted from leaves and was rich in linalool components as found in lavender and jasmine oil. -
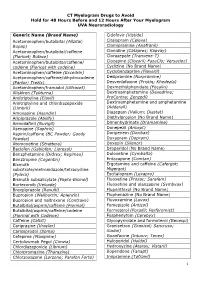
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology
CT Myelogram Drugs to Avoid Hold for 48 Hours Before and 12 Hours After Your Myelogram UVA Neuroradiology Generic Name (Brand Name) Cidofovir (Vistide) Acetaminophen/butalbital (Allzital; Citalopram (Celexa) Bupap) Clomipramine (Anafranil) Acetaminophen/butalbital/caffeine Clonidine (Catapres; Kapvay) (Fioricet; Butace) Clorazepate (Tranxene-T) Acetaminophen/butalbital/caffeine/ Clozapine (Clozaril; FazaClo; Versacloz) codeine (Fioricet with codeine) Cyclizine (No Brand Name) Acetaminophen/caffeine (Excedrin) Cyclobenzaprine (Flexeril) Acetaminophen/caffeine/dihydrocodeine Desipramine (Norpramine) (Panlor; Trezix) Desvenlafaxine (Pristiq; Khedezla) Acetaminophen/tramadol (Ultracet) Dexmethylphenidate (Focalin) Aliskiren (Tekturna) Dextroamphetamine (Dexedrine; Amitriptyline (Elavil) ProCentra; Zenzedi) Amitriptyline and chlordiazepoxide Dextroamphetamine and amphetamine (Limbril) (Adderall) Amoxapine (Asendin) Diazepam (Valium; Diastat) Aripiprazole (Abilify) Diethylpropion (No Brand Name) Armodafinil (Nuvigil) Dimenhydrinate (Dramamine) Asenapine (Saphris) Donepezil (Aricept) Aspirin/caffeine (BC Powder; Goody Doripenem (Doribax) Powder) Doxapram (Dopram) Atomoxetine (Strattera) Doxepin (Silenor) Baclofen (Gablofen; Lioresal) Droperidol (No Brand Name) Benzphetamine (Didrex; Regimex) Duloxetine (Cymbalta) Benztropine (Cogentin) Entacapone (Comtan) Bismuth Ergotamine and caffeine (Cafergot; subcitrate/metronidazole/tetracycline Migergot) (Pylera) Escitalopram (Lexapro) Bismuth subsalicylate (Pepto-Bismol) Fluoxetine (Prozac; Sarafem) -

Tramadol 50 Mg Capsules Tramadol Hydrochloride
Tramadol 50 mg Capsules Tramadol hydrochloride PACKAGE LEAFLET: INFORMATION FOR THE USER Read all of this leaflet carefully before you start taking this medicine. • Keep this leaflet. You may need to read it again. • If you have any further questions, ask your doctor or pharmacist. • This medicine has been prescribed for you. Do not pass it on to others. It may harm them, even if their symptoms are the same as yours. • If any of the side effects gets serious, or if you notice any side effects not listed in this leaflet, please tell your doctor or pharmacist. In this leaflet 1. What are Tramadol 50 mg Capsules and what are they used for 2. What you need to know before you take Tramadol 50 mg Capsules 3. How to take Tramadol 50 mg Capsules 4. Possible side effects 5. How to store Tramadol 50 mg Capsules 6. Contents of the pack and other information 1. WHAT ARE TRAMADOL 50 mg CAPSULES AND WHAT ARE THEY USED FOR Tramadol 50 mg Capsules are 'analgesics' which act on the central nervous system (the brain and the spinal cord). Analgesics are often called 'pain killers' or 'pain relievers'. Tramadol 50 mg Capsules relieve pain and can also be taken to prevent pain. Pain is a symptom not an illness. There are many types of pain with many different causes, for example back- ache, toothache, pain after an operation or pain from broken bones. Tramadol 50 mg Capsules help your body's system for relieving pain. It does this in two ways: - Acts directly on parts of your brain and spinal cord to reduce the amount of pain you feel - Reduces the size of the pain message passed from one nerve to another.