Araneae: Pholcidae) Affects Whirling and Other Defensive Behaviors
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Checklist of the Non -Acarine Arachnids
Original Research A CHECKLIST OF THE NON -A C A RINE A R A CHNIDS (CHELICER A T A : AR A CHNID A ) OF THE DE HOOP NA TURE RESERVE , WESTERN CA PE PROVINCE , SOUTH AFRIC A Authors: ABSTRACT Charles R. Haddad1 As part of the South African National Survey of Arachnida (SANSA) in conserved areas, arachnids Ansie S. Dippenaar- were collected in the De Hoop Nature Reserve in the Western Cape Province, South Africa. The Schoeman2 survey was carried out between 1999 and 2007, and consisted of five intensive surveys between Affiliations: two and 12 days in duration. Arachnids were sampled in five broad habitat types, namely fynbos, 1Department of Zoology & wetlands, i.e. De Hoop Vlei, Eucalyptus plantations at Potberg and Cupido’s Kraal, coastal dunes Entomology University of near Koppie Alleen and the intertidal zone at Koppie Alleen. A total of 274 species representing the Free State, five orders, 65 families and 191 determined genera were collected, of which spiders (Araneae) South Africa were the dominant taxon (252 spp., 174 genera, 53 families). The most species rich families collected were the Salticidae (32 spp.), Thomisidae (26 spp.), Gnaphosidae (21 spp.), Araneidae (18 2 Biosystematics: spp.), Theridiidae (16 spp.) and Corinnidae (15 spp.). Notes are provided on the most commonly Arachnology collected arachnids in each habitat. ARC - Plant Protection Research Institute Conservation implications: This study provides valuable baseline data on arachnids conserved South Africa in De Hoop Nature Reserve, which can be used for future assessments of habitat transformation, 2Department of Zoology & alien invasive species and climate change on arachnid biodiversity. -

Untangling Taxonomy: a DNA Barcode Reference Library for Canadian Spiders
Molecular Ecology Resources (2016) 16, 325–341 doi: 10.1111/1755-0998.12444 Untangling taxonomy: a DNA barcode reference library for Canadian spiders GERGIN A. BLAGOEV, JEREMY R. DEWAARD, SUJEEVAN RATNASINGHAM, STEPHANIE L. DEWAARD, LIUQIONG LU, JAMES ROBERTSON, ANGELA C. TELFER and PAUL D. N. HEBERT Biodiversity Institute of Ontario, University of Guelph, Guelph, ON, Canada Abstract Approximately 1460 species of spiders have been reported from Canada, 3% of the global fauna. This study provides a DNA barcode reference library for 1018 of these species based upon the analysis of more than 30 000 specimens. The sequence results show a clear barcode gap in most cases with a mean intraspecific divergence of 0.78% vs. a min- imum nearest-neighbour (NN) distance averaging 7.85%. The sequences were assigned to 1359 Barcode index num- bers (BINs) with 1344 of these BINs composed of specimens belonging to a single currently recognized species. There was a perfect correspondence between BIN membership and a known species in 795 cases, while another 197 species were assigned to two or more BINs (556 in total). A few other species (26) were involved in BIN merges or in a combination of merges and splits. There was only a weak relationship between the number of specimens analysed for a species and its BIN count. However, three species were clear outliers with their specimens being placed in 11– 22 BINs. Although all BIN splits need further study to clarify the taxonomic status of the entities involved, DNA bar- codes discriminated 98% of the 1018 species. The present survey conservatively revealed 16 species new to science, 52 species new to Canada and major range extensions for 426 species. -

The Pholcid Spiders of Micronesia and Polynesia (Araneae, Pholcidae) Joseph A
Butler University Digital Commons @ Butler University Scholarship and Professional Work - LAS College of Liberal Arts & Sciences 2008 The pholcid spiders of Micronesia and Polynesia (Araneae, Pholcidae) Joseph A. Beatty James W. Berry Butler University, [email protected] Bernhard A. Huber Follow this and additional works at: http://digitalcommons.butler.edu/facsch_papers Part of the Biology Commons, and the Entomology Commons Recommended Citation Beatty, Joseph A.; Berry, James W.; and Huber, Bernhard A., "The hop lcid spiders of Micronesia and Polynesia (Araneae, Pholcidae)" Journal of Arachnology / (2008): 1-25. Available at http://digitalcommons.butler.edu/facsch_papers/782 This Article is brought to you for free and open access by the College of Liberal Arts & Sciences at Digital Commons @ Butler University. It has been accepted for inclusion in Scholarship and Professional Work - LAS by an authorized administrator of Digital Commons @ Butler University. For more information, please contact [email protected]. The pholcid spiders of Micronesia and Polynesia (Araneae, Pholcidae) Author(s): Joseph A. Beatty, James W. Berry, Bernhard A. Huber Source: Journal of Arachnology, 36(1):1-25. Published By: American Arachnological Society DOI: http://dx.doi.org/10.1636/H05-66.1 URL: http://www.bioone.org/doi/full/10.1636/H05-66.1 BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use. -

Arachnida, Solifugae) with Special Focus on Functional Analyses and Phylogenetic Interpretations
HISTOLOGY AND ULTRASTRUCTURE OF SOLIFUGES Comparative studies of organ systems of solifuges (Arachnida, Solifugae) with special focus on functional analyses and phylogenetic interpretations HISTOLOGIE UND ULTRASTRUKTUR DER SOLIFUGEN Vergleichende Studien an Organsystemen der Solifugen (Arachnida, Solifugae) mit Schwerpunkt auf funktionellen Analysen und phylogenetischen Interpretationen I N A U G U R A L D I S S E R T A T I O N zur Erlangung des akademischen Grades doctor rerum naturalium (Dr. rer. nat.) an der Mathematisch-Naturwissenschaftlichen Fakultät der Ernst-Moritz-Arndt-Universität Greifswald vorgelegt von Anja Elisabeth Klann geboren am 28.November 1976 in Bremen Greifswald, den 04.06.2009 Dekan ........................................................................................................Prof. Dr. Klaus Fesser Prof. Dr. Dr. h.c. Gerd Alberti Erster Gutachter .......................................................................................... Zweiter Gutachter ........................................................................................Prof. Dr. Romano Dallai Tag der Promotion ........................................................................................15.09.2009 Content Summary ..........................................................................................1 Zusammenfassung ..........................................................................5 Acknowledgments ..........................................................................9 1. Introduction ............................................................................ -

Records of the Hawaii Biological Survey for 1996
Records of the Hawaii Biological Survey for 1996. Bishop Museum Occasional Papers 49, 71 p. (1997) RECORDS OF THE HAWAII BIOLOGICAL SURVEY FOR 1996 Part 2: Notes1 This is the second of 2 parts to the Records of the Hawaii Biological Survey for 1996 and contains the notes on Hawaiian species of protists, fungi, plants, and animals includ- ing new state and island records, range extensions, and other information. Larger, more comprehensive treatments and papers describing new taxa are treated in the first part of this Records [Bishop Museum Occasional Papers 48]. Foraminifera of Hawaii: Literature Survey THOMAS A. BURCH & BEATRICE L. BURCH (Research Associates in Zoology, Hawaii Biological Survey, Bishop Museum, 1525 Bernice Street, Honolulu, HI 96817, USA) The result of a compilation of a checklist of Foraminifera of the Hawaiian Islands is a list of 755 taxa reported in the literature below. The entire list is planned to be published as a Bishop Museum Technical Report. This list also includes other names that have been applied to Hawaiian foraminiferans. Loeblich & Tappan (1994) and Jones (1994) dis- agree about which names should be used; therefore, each is cross referenced to the other. Literature Cited Bagg, R.M., Jr. 1980. Foraminifera collected near the Hawaiian Islands by the Steamer Albatross in 1902. Proc. U.S. Natl. Mus. 34(1603): 113–73. Barker, R.W. 1960. Taxonomic notes on the species figured by H. B. Brady in his report on the Foraminifera dredged by HMS Challenger during the years 1873–1876. Soc. Econ. Paleontol. Mineral. Spec. Publ. 9, 239 p. Belford, D.J. -

Zootaxa, a Review of the Cellar Spider Genus Psilochorus Simon 1893 In
Zootaxa 2144: 1–53 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) A review of the cellar spider genus Psilochorus Simon 1893 in America north of Mexico (Araneae: Pholcidae) JOZEF SLOWIK University of Alaska Museum of the North, Department of Entomology, 907 Yukon Drive, Fairbanks, Alaska 99775 USA. E-mail: [email protected] Table of contents Abstract ............................................................................................................................................................................... 2 Introduction ......................................................................................................................................................................... 2 Materials and methods ........................................................................................................................................................ 2 Taxonomy ............................................................................................................................................................................ 3 Pholcidae C. L. Koch, 1851 ................................................................................................................................................ 3 Psilochorus Simon, 1893 .................................................................................................................................................... 3 Key to Psilochorus species in America -

Circadian Rhythms of the Spider Pholcus Phalangeoides in Activity Monitors and Web Boxes
East Tennessee State University Digital Commons @ East Tennessee State University Undergraduate Honors Theses Student Works 5-2019 Circadian Rhythms of the Spider Pholcus phalangeoides in Activity Monitors and Web Boxes Steven Dirmeyer Follow this and additional works at: https://dc.etsu.edu/honors Part of the Animal Experimentation and Research Commons, Behavior and Ethology Commons, and the Biology Commons Recommended Citation Dirmeyer, Steven, "Circadian Rhythms of the Spider Pholcus phalangeoides in Activity Monitors and Web Boxes" (2019). Undergraduate Honors Theses. Paper 640. https://dc.etsu.edu/honors/640 This Honors Thesis - Open Access is brought to you for free and open access by the Student Works at Digital Commons @ East Tennessee State University. It has been accepted for inclusion in Undergraduate Honors Theses by an authorized administrator of Digital Commons @ East Tennessee State University. For more information, please contact [email protected]. Circadian Rhythms of the Spider Pholcus phalangeoides in Activity Monitors and Web Boxes Thesis submitted in partial fulfillment of Honors By Steven Dirmeyer The Honors College University Honors Scholars Program East Tennessee State University April (26), 2019 --------------------------------------------- Dr. Thomas C. Jones, Faculty Mentor --------------------------------------------- Dr. Darrell J. Moore, Faculty Reader SPIDER CIRCADIAN RHYTHMS IN ACTIVITY MONITORS AND WEB BOXES 1 Abstract: Circadian rhythms are endogenous molecular clocks that correspond to the 24-hour day and are regulated by light stimulus, allowing organisms to entrain to the dawn-dusk cycle. These clocks may allow organisms to anticipate daily events, influencing their behavior. In arthropods, including spiders, circadian rhythmicity is tested using activity monitors, which house individuals in tubes. However, this does not reflect the natural habitat of many spiders. -

Arthropods of Elm Fork Preserve
Arthropods of Elm Fork Preserve Arthropods are characterized by having jointed limbs and exoskeletons. They include a diverse assortment of creatures: Insects, spiders, crustaceans (crayfish, crabs, pill bugs), centipedes and millipedes among others. Column Headings Scientific Name: The phenomenal diversity of arthropods, creates numerous difficulties in the determination of species. Positive identification is often achieved only by specialists using obscure monographs to ‘key out’ a species by examining microscopic differences in anatomy. For our purposes in this survey of the fauna, classification at a lower level of resolution still yields valuable information. For instance, knowing that ant lions belong to the Family, Myrmeleontidae, allows us to quickly look them up on the Internet and be confident we are not being fooled by a common name that may also apply to some other, unrelated something. With the Family name firmly in hand, we may explore the natural history of ant lions without needing to know exactly which species we are viewing. In some instances identification is only readily available at an even higher ranking such as Class. Millipedes are in the Class Diplopoda. There are many Orders (O) of millipedes and they are not easily differentiated so this entry is best left at the rank of Class. A great deal of taxonomic reorganization has been occurring lately with advances in DNA analysis pointing out underlying connections and differences that were previously unrealized. For this reason, all other rankings aside from Family, Genus and Species have been omitted from the interior of the tables since many of these ranks are in a state of flux. -
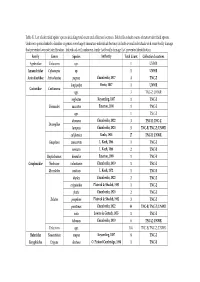
Table S1. List of Identified Spider Species Including Total Count and Collection Locations. Bold Cells Include Counts of Mature Identified Species
Table S1. List of identified spider species including total count and collection locations. Bold cells include counts of mature identified species. Unknown species linked to families or genera were largely immature individuals but may include several individuals with some bodily damage that prevented accurate identification. Individuals with unknown family had bodily damage that prevented identification. Family Genus Species Authority Total Count Collection Locations Agelenidae Unknown spp. 1 UNWR Amaurobiidae Cybaeopsis sp. 1 UNWR Antrodiaetidae Antrodiaetus pugnax Chamberlin, 1917 4 TNC-Z longipalpa Hentz, 1847 1 UNWR Corinnidae Castianeira spp. 3 TNC-Z; UNWR neglectus Keyserling, 1887 1 TNC-Z Drassodes saccatus Emerton, 1890 1 TNC-Z spp. 1 TNC-Z dromeus Chamberlin, 1922 3 TNC-B; TNC-Z Drassyllus lamprus Chamberlin, 1920 3 TNC-B; TNC-Z; UNWR californica Banks, 1904 17 TNC-B; UNWR Gnaphosa muscorum L. Koch, 1866 1 TNC-Z sericata L. Koch, 1866 2 TNC-B Haplodrassus hiemalis Emerton, 1909 1 TNC-B Gnaphosidae Nodocion voluntaries Chamberlin, 1919 1 TNC-Z Urozelotes rusticus L. Koch, 1872 1 TNC-B duplex Chamberlin, 1922 2 TNC-Z exiguioides Platnick & Shadab, 1983 1 TNC-Z fratis Chamberlin, 1920 2 TNC-Z Zelotes josephine Platnick & Shadab, 1983 3 TNC-Z puritanus Chamberlin, 1922 44 TNC-B; TNC-Z; UNWR sula Lowrie & Gertsch, 1955 1 TNC-Z tubuous Chamberlin, 1919 6 TNC-Z; UNWR Unknown spp. 184 TNC-B; TNC-Z; UNWR Hahniidae Neoantistea magna Keyserling, 1887 8 TNC-Z Linyphiidae Erigone dentosa O. Pickard-Cambridge, 1894 1 TNC-B spp. 1 TNC-Z Unknown spp. 13 TNC-Z; UNWR mccooki Montgomery, 1904 95 TNC-B; TNC-Z; UNWR Schizocosa minnesotensis Gertsch, 1934 25 TNC-B Lycosidae spp. -

3. Zoo.DIVERSITY
International Journal of Zoology and Research (IJZR) ISSN(P): 2278-8816; ISSN(E): 2278-8824 Vol. 6, Issue 1, Feb 2016, 15-24 © TJPRC Pvt. Ltd. EXPLORATIVE STUDY ON THE DIVERSITY AND CHARACTERISTICS OF SPIDER FAMILIES VARSHA W WANKHADE & NARENDRA MANWAR Department of Zoology, University of Pune, Pune, Maharashtra, India ABSTRACT The order Aranae deserves a special place in the study of ecology as it acts as a biological indicator. The objectives of the present study were to explore the diversity and characteristics of spider families. The study was conducted at Sawanga-Vithoba lake (Malkhed Project) district Amravati, Maharashtra India. In this study, by observing the diversity of Spiders (Class Arachnida, phylum Arthropoda), efforts were made to evaluate the status of ecosystem of Sawanga-Vithoba Lake (Malkhed Project) district Amravati, Maharashtra. In total 42 species of spiders belonging to 14 families were observed. Araneidae was the most represented family with 15 species. Area surrounding Sawanga-Vithoba lake (Malkhed Project) Dist Amravati, Maharashtra, India represents 23.72 % of the total families recorded in India. The high species diversity of spiders in Sawanga-Vithoba lake region can be attributed to the high diversity of plants and insects. In totality, five different guilds of spiders were observed. 54% of spiders were orb web builders, 24% were foliage runner. Ground runner and Ambusers were 8%, while scattered line weaver were 6%. Sawanga-Vithoba lake region Original Article Article Original could be an important centre of speciation in Pohara forest of tahsil chandur railway dist Amravati. This is the first report of the spider fauna from Sawanga-Vithoba lake region. -

Note on Suspected Brown Recluse Spiders (Araneae: Sicariidae) in South Carolina
Faculty Research Note Note on Suspected Brown Recluse Spiders (Araneae: Sicariidae) in South Carolina Robert J. Wolff* South University, 9 Science Court, Columbia, SC 29203 The general public believes that brown recluse spiders (Loxosceles Filistatidae (Kukulcania hibernalis) 22 specimens reclusa) are widespread where they live and that these spiders are Lycosidae 21 (3 in one package, 5 in another) frequent causes of bites resulting in dermonecrosis. Research over the Pholcidae 17 past twenty years shows these reports to be unfounded. Vetter (2005) Miturgidae 8 examined 1,773 specimens sent in from across the U.S. as brown recluse Theridiidae 8 spiders and no specimens were found from areas outside the species Agelenidae 7 range, with the exception of a specimen from California. Araneidae 6 Clubionidae 6 The reported range of the brown recluse spider includes all or major Thomisidae 6 portions of Arkansas, Oklahoma, Texas, Louisiana, Alabama, Tennessee, Gnaphosidae 4 Kentucky, Illinois, Missouri and Kansas. Minor portions of the brown Corinnidae 3 recluse range were previously reported in Iowa, Indiana, Ohio, New Philodromidae 3 Mexico, North Carolina, Georgia, and South Carolina. The most recent Amaurobiidae 1 map (Vetter, 2015) does not include South Carolina, and only the far Pisauridae 1 western tip of North Carolina and northwestern corner of Georgia. Scytodidae (Scytodes thoracica) 1 Unidentifiable 4 Schuman and Caldwell (1991) found that South Carolina physicians reported treating 478 cases of brown recluse spider envenomations in 1990 alone. This seems like a very high number, unfortunately all or No brown recluses were identified from the specimens obtained in this almost all of these are probably not brown recluse spider bites. -

Pest Profile
Pest Profile Photo credit: Gary Alpert, Harvard University, Bugwood.org Licensed under a Creative Commons Attribution-Noncommercial 3.0 License Common Name: Long-bodied Cellar spider Scientific Name: Pholcus phalangioides Order and Family: Opiliones; Phalangida Size and Appearance: Length (mm) Appearance Egg - Laid in clusters of 25-60 1mm - Wrapped in a layer of silk - Carried by female Larva/Nymph - Have a small, gray or brown body 1-2mm - Long legs Adult - Have a small, cylindrical, and elongated body - Grayish-brown with either translucent areas or patches of dark gray and beige Body: 6-10 mm - Very long, thin grayish-brown legs, covered in Legs: 55 mm tiny hairs Pupa (if applicable) Type of feeder (Chewing, sucking, etc.): Chelicerae for piercing and chewing Host plant/s: Found in and/or around man-made structures or in other undisturbed, poorly lit habitats Description of Damage (larvae and adults): Because these spiders tend to live inside man-made structures such as house, their webs and presence make them a pest. References: Ferrick, A. (2002). Pholcus phalangioides. Retrieved September 27, 2017, from http://animaldiversity.org/accounts/Pholcus_phalangioides/ Boone, M., Elliott, L., Heins, C., Barnd, B., Entz, C., Schneider, K., & Howe, M. (2005, January 08). Species Pholcus phalangioides - Longbodied Cellar Spider. Retrieved September 25, 2017, from http://bugguide.net/node/view/9610 Department of Entomology - Penn State College of Agricultural Sciences. (2002, March). Longbodied Cellar Spider (Department of Entomology). Retrieved September 27, 2017, from http://ento.psu.edu/extension/factsheets/longbodied-cellar-spider Spiders.us. (2016, May 28). Pholcus phalangioides (Longbodied Cellar Spider). Retrieved December 6, 2016, from http://www.spiders.us/species/pholcus-phalangioides/ .