Uvic Thesis Template
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Rockfish Population Slowly on the Rise
West Coast rockfish Rockfish population slowly on the rise Above: Rockfish in a kelp forest. Right: Black rockfish. Inset: Juvenile rockfish. More than 60 species of rockfi sh dwell in ocean perch, and canary. These species are of- schooling rockfi sh. vast numbers in the Eastern Pacifi c Ocean fi cially “overfi shed,” a designation which the When you’ve got a permit for hake, you’re from Baja to Alaska. White-fl eshed and deli- council grants when a fi sh’s population drops geared up for hake, you’re fi shing for hake, cious, they frequently school together and to 25 percent of its estimated virgin level, and and you know there is a hell of a lot of hake often intermingle with other commercially a stock is not considered “recovered” until it around, nothing stinks so much as scooping sought fi shes — but seven members of the ge- has climbed back up to the 40-percent mark. up a school of lousy rockfi sh — especially nus Sebastes have spoiled the party. These guidelines were devised and written ones protected by strict bycatch limits. Actually, it was fi shermen, who harvested into policy in 1998 as part of the Fishery Man- The fl eet may have dozens of tonnes of hake them nearly to economic extinction last centu- agement Plan. Since then, two species have as- left uncaught in the quota but must stop be- ry, and this handful of species is now largely cended back to the 40 percent level and been cause the rockfi sh cap has been exceeded. -

Vii Fishery-At-A-Glance: California Sheephead
Fishery-at-a-Glance: California Sheephead (Sheephead) Scientific Name: Semicossyphus pulcher Range: Sheephead range from the Gulf of California to Monterey Bay, California, although they are uncommon north of Point Conception. Habitat: Both adult and juvenile Sheephead primarily reside in kelp forest and rocky reef habitats. Size (length and weight): Male Sheephead can grow up to a length of three feet (91 centimeters) and weigh over 36 pounds (16 kilograms). Life span: The oldest Sheephead ever reported was a male at 53 years. Reproduction: As protogynous hermaphrodites, all Sheephead begin life as females, and older, larger females can develop into males. They are batch spawners, releasing eggs and sperm into the water column multiple times during their spawning season from July to September. Prey: Sheephead are generalist carnivores whose diet shifts throughout their growth. Juveniles primarily consume small invertebrates like tube-dwelling polychaetes, bryozoans and brittle stars, and adults shift to consuming larger mobile invertebrates like sea urchins. Predators: Predators of adult Sheephead include Giant Sea Bass, Soupfin Sharks and California Sea Lions. Fishery: Sheephead support both a popular recreational and a commercial fishery in southern California. Area fished: Sheephead are fished primarily south of Point Conception (Santa Barbara County) in nearshore waters around the offshore islands and along the mainland shore over rocky reefs and in kelp forests. Fishing season: Recreational anglers can fish for Sheephead from March 1through December 31 onboard boats south of Point Conception and May 1 through December 31 between Pigeon Point and Point Conception. Recreational divers and shore-based anglers can fish for Sheephead year round. -

WCR-2018-10181 October 4, 2018
UNITED STATES DEPARTMENT OF COMMERCE National Oceanic and Atmospheric Administration NATIONAL MARINE FISHERIES SERVICE West Coast Region 1201 NE Lloyd Boulevard, Suite 1100 Portland, OR 97232 Refer to NMFS No.: WCR-2018-10181 October 4, 2018 Amy Changchien Director, Office of Planning and Program Development U.S. Department of Transportation Federal Transit Administration 915 Second Avenue Federal Bldg. Suite 3142 Seattle, Washington 98174-1002 Re: Reinitiation of Endangered Species Act Section 7(a)(2) Biological Opinion, and Magnuson-Stevens Fishery Conservation and Management Act Essential Fish Habitat Response for One Structure in Inland Marine Waters of Washington State: Annapolis Ferry Terminal Pier, Ramp, and Float Structure. Dear Ms. Changchien: Thank you for your June 6, 2018, letter requesting reinitiation of consultation with NOAA’s National Marine Fisheries Service (NMFS) pursuant to section 7 of the Endangered Species Act of 1973 (ESA) (16 U.S.C. 1531 et seq.). We also completed an essential fish habitat (EFH) consultation on the proposed action, in accordance with section 305(b)(2) of the Magnuson- Stevens Fishery Conservation and Management Act (MSA) (16 U.S.C. 1801 et seq.) and implementing regulations at 50 CFR 600. The enclosed document contains a biological opinion (opinion) that analyzes the effects of your proposal to permit the removal of one pier, ramp, float (PRF) structure and the installation of one PRF structure. In this opinion, we conclude that the proposed action is not likely to jeopardize the continued existence of Puget Sound (PS) Chinook salmon (Oncorhynchus tshawytscha), PS steelhead (O. mykiss), or PS/Georgia Basin (GB) bocaccio (Sebastes paucispinis), PS/GB yelloweye rockfish (S. -

Rockfish Resources of the South Central California Coast
140-155 Stephens 11/13/06 9:02 PM Page 140 STEPHENS ET AL.: ROCKFISH RESOURCES OF SOUTH CENTRAL CA CalCOFI Rep., Vol. 47, 2006 ROCKFISH RESOURCES OF THE SOUTH CENTRAL CALIFORNIA COAST: ANALYSIS OF THE RESOURCE FROM PARTYBOAT DATA, 1980–2005 JOHN STEPHENS DEAN WENDT DEBRA WILSON-VANDENBERG California Polytechnic State University California Polytechnic State University, California Department of Fish and Game San Luis Obispo and Vantuna Research Group San Luis Obispo 20 Lower Ragsdale Drive 2550 Nightshade Place 2550 Nightshade Place Monterey, California 93940 Arroyo Grande, California 93420 Arroyo Grande, California 93420 [email protected] JAY CARROLL ROYDEN NAKAMURA ERIN NAKADA Tenera Environmental, Inc. Biological Sciences Biological Sciences 141 Suburban Road, Suite A2 California Polytechnic State University California Polytechnic State University San Luis Obispo, California 93401 San Luis Obispo, California 93407 San Luis Obispo, California 93407 STEVEN RIENECKE JONO WILSON Biological Sciences Biological Sciences California Polytechnic State University California Polytechnic State University San Luis Obispo, California 93407 San Luis Obispo, California 93407 ABSTRACT including Lenarz (1987), Ralston (1998), Gunderson Rockfishes (Sebastes spp.) have historically comprised (1998), and Love et al. (1998, 2002). Rockfish are long- a large proportion of catches in the nearshore recre lived, slow to mature (iteroparous), and therefore sub ational fishery in California, but declining populations ject to pre-spawning mortality (Leaman 1991). Two of some species have led to increasingly restrictive man factors, overfishing and climate change, are considered agement of the resource. This report summarizes new primarily responsible for the declining marine fish pop and existing data on rockfishes of the south central coast ulations in much of California. -
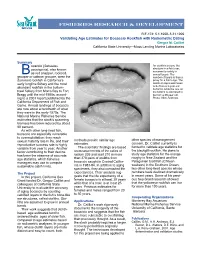
Validating Age Estimates for Bocaccio Rockfish with Radiometric Dating Gregor M
FISHERIES RESEARCH & DEVELOPMENT R/F-174: 5.1.1998–5.31.1999 Validating Age Estimates for Bocaccio Rockfish with Radiometric Dating Gregor M. Cailliet California State University—Moss Landing Marine Laboratories Summary ocaccio (Sebastes An otolith is a bone-like paucispinis), also known structure in a fish’s ear, assumed to calcify in Bas red snapper, rockcod, annual layers. The grouper or salmon grouper, were the number of layers is thus a dominant rockfish in California’s proxy for a fish's age. The early longline fishery and the most otolith on top is estimated to be from a 3-year-old abundant rockfish in the bottom- bocaccio while the one on trawl fishery from Morro Bay to Fort the bottom is estimated to Bragg until the mid-1980s, accord- be from a 28-year-old. ing to a 2001 report published by the Photo: Allen Andrews. California Department of Fish and Game. Annual landings of bocaccio are now about a hundredth of what they were in the early-1970s. The National Marine Fisheries Service estimates that the stock’s spawning biomass has been reduced by about 98 percent. As with other long-lived fish, bocaccio are especially vulnerable to overexploitation: they reach sexual maturity late in life, and their methods provide similar age other species of management reproductive success rate is highly estimates. concern. Dr. Cailliet currently is variable from year to year. Another The scientists’ findings are based funded to validate age statistics for factor contributing to their decline on measurements of the ratios of the blackgill rockfish. -

The Distribution of Pelagic Juvenile Rockfish of the Genus Sebastes in the Upwelling Region Off Central California Ralph J
LARSON ET AL.: DISTRIBUTION OF PELAGIC JUVENILE ROCKFISH CalCOFl Rep., Vol. 35, 1994 THE DISTRIBUTION OF PELAGIC JUVENILE ROCKFISH OF THE GENUS SEBASTES IN THE UPWELLING REGION OFF CENTRAL CALIFORNIA RALPH J. LARSON WILLIAM H. LENARZ AND STEPHEN RALSTON Department of Biology National Manne Fishenes Semce San Francisco State University 31 50 Paradlse Dnve San Francisco. California 94132 Tiburon, California 94920 ABSTRACT en la composici6n por especies y tallas. Los juveniles Many species of central California rockfishes conclude peligicos mas pequeiios se encontraron a menudo ale- their pelagic stage during the spring-summer upwelling jados de la costa, en la regibn asociada a1 frente de la sur- period, when advection of surface waters could carry gencia, lo que sugiere que fueron transportados por la them away from nearshore postpelagic habitats. We ex- corriente de surgencia. Sin embargo, pocos juveniles amined the distributions of late-stage pelagic juvenile peligicos fueron encontrados del lado hacia mar aden- rockfish in April and May/June of 1987 and 1988, based tro del frente de la surgencia. Los juveniles peligicos de on midwater-trawl surveys between Point Reyes and mayor tamaiio se encontraron a menudo pr6ximos a la Monterey Bay, California. Distributional patterns were costa, aun en 10s periodos de actividad de la surgencia. complex, and changed rapidly with changing oceano- Lo anterior sugiere que 10s juveniles peligicos cambian graphic conditions and with changes in species and size su comportamiento durante sus fases tardias, lo que les composition. The smallest pelagic juveniles often ap- permite desplazarse hacia la costa, o bien, permanecer peared offshore, in the region of the upwelling front, alli a pesar de la surgencia. -

Zalophus Californianus): Insights from Molecular Scatology
San Jose State University SJSU ScholarWorks Master's Theses Master's Theses and Graduate Research Fall 2016 Sex-specific diet and ockfishr consumption in California sea lions (Zalophus californianus): Insights from molecular scatology Keith Michael Hernandez San Jose State University Follow this and additional works at: https://scholarworks.sjsu.edu/etd_theses Recommended Citation Hernandez, Keith Michael, "Sex-specific diet and ockfishr consumption in California sea lions (Zalophus californianus): Insights from molecular scatology" (2016). Master's Theses. 4757. DOI: https://doi.org/10.31979/etd.49wc-7qbn https://scholarworks.sjsu.edu/etd_theses/4757 This Thesis is brought to you for free and open access by the Master's Theses and Graduate Research at SJSU ScholarWorks. It has been accepted for inclusion in Master's Theses by an authorized administrator of SJSU ScholarWorks. For more information, please contact [email protected]. SEX-SPECIFIC DIET AND ROCKFISH CONSUMPTION IN CALIFORNIA SEA LIONS ( ZALOPHUS CALIFORNIANUS ): INSIGHTS FROM MOLECULAR SCATOLOGY A Thesis Presented to The Faculty of Moss Landing Marine Laboratories San José State University In Partial Fulfillment of the Requirements for the Degree Master of Science by Keith M. Hernandez December 2016 © 2016 Keith Michael Hernandez ALL RIGHTS RESERVED The Designated Thesis Committee Approves the Thesis Titled SEX-SPECIFIC DIET AND ROCKFISH CONSUMPTION IN CALIFORNIA SEA LIONS ( ZALOPHUS CALIFORNIANUS ): INSIGHTS FROM MOLECULAR SCATOLOGY by Keith M. Hernandez APPROVED FOR THE DEPARTMENT OF MARINE SCIENCE SAN JOSÉ STATE UNIVERSITY December 2016 Dr. James T. Harvey Moss Landing Marine Laboratories Dr. Jonathan B. Geller Moss Landing Marine Laboratories Dr. John Carlos Garza National Oceanic and Atmospheric Administration, Southwest Fisheries Science Center ABSTRACT SEX-SPECIFIC DIET AND ROCKFISH CONSUMPTION IN CALIFORNIA SEA LIONS ( ZALOPHUS CALIFORNIANUS ): INSIGHTS FROM MOLECULAR SCATOLOGY By Keith M. -

Appendix B Species Biology
APPENDIX B SPECIES BIOLOGY Appendix B SPECIES BIOLOGY Species covered in this Biological Assessment Reference were chosen on the basis of species lists obtained from the National Marine Fisheries Service (NMFS) and the U.S. Fish and Wildlife Service (USFWS) (see Appendix C). Because of the inland marine nature of all the Washington State Ferries (WSF) terminals, freshwater fish, oceangoing mammals, and sea turtles (including leatherback sea turtles), terrestrial species, and upland and freshwater plants were excluded from this analysis. NMFS – Listed Threatened, Endangered, and Proposed Species Puget Sound Chinook Salmon (Oncorhynchus tshawytscha) Status Puget Sound Chinook salmon were listed as threatened on August 2, 1999 (Federal Register 1999a.). The evolutionarily significant unit (ESU) includes naturally spawned fish and stocks from 26 artificial propagation programs. Critical Habitat Critical habitat was designated for Puget Sound Chinook salmon on February 16, 2000 (Federal Register 2000.). Critical habitat is designated for areas containing the physical and biological habitat features, or primary constituent elements (PCEs) essential for the conservation of the species or which require special management considerations. PCEs include sites that are essential to supporting one or more life stages of the ESU and that contain physical or biological features essential to the conservation of the ESU. Specific sites and features designated for Puget Sound Chinook salmon include the following: 1. Freshwater spawning sites with water quantity and quality conditions and substrate supporting spawning incubation and larval development 2. Freshwater rearing sites with water quantity and floodplain connectivity to form and maintain physical habitat conditions and support juvenile growth and mobility; water quality and forage supporting juvenile development; and natural cover such as shade, submerged and overhanging large wood, log jams and beaver dams, aquatic vegetation, large rocks and boulders, side channels, and undercut banks 3. -

NOAA/NWFSC Southern California Shelf Rockfish Hook and Line Survey
NOAA/NWFSC Southern California Shelf Rockfish Hook and Line Survey NOAA Fisheries, Northwest Fisheries Science Center 2725 Montlake Blvd. E, Seattle, WA 98112 John Harms, NOAA/NWFSC Jim Benante, PSMFC/NWFSC Matt Barnhart, PSMFC/NWFSC Rationale • Prior to 2002, there was no ongoing fishery- independent groundfish data collection for the area from Pt. Conception (34 27΄ N) to the Mexican border) • Trawl survey coverage for the region began in 2002; however, untrawlable areas including rocky, high-relief habitats remained unsampled Survey Region: Southern California Bight Primary Objective Develop an annual index of abundance and time series of other biological parameters for key species of structure-associated shelf rockfish (Genus: Sebastes) within the Southern California Bight including bocaccio, vermilion rockfish, sunset rockfish, greenspotted rockfish, and others. Background • Initial planning: Summer 2002 – Meetings between NOAA Fisheries researchers and representatives from the sportfishing and commercial fishing industry • Pilot survey: May 2003 – Research conducted aboard vessels chartered from both the sportfishing and commercial industries • Survey has been conducted annually since 2004 – 2011 will be the 8th year in the data time series Survey Design • Conducted annually • 121 fixed stations sampled during the course of the survey • Sampling area: Pt. Arguello to the boundary with Mexican waters • Depth range: 40m – 230m (common depth range for bocaccio) • Sampling frame developed through meetings with the local sport and commercial fishermen; previous groundfish monitoring programs conducted by California Dept. of Fish and Game; and stations sampled opportunistically during the 2003 pilot study Survey Stations Point Conception Santa Barbara Santa Barbara Channel Point Hueneme San Harrison Reef Miguel Is. -

Summarized History of Oregon Marine Sport Fishing Regulations
Summarized History of Oregon Marine Sport Fishing Regulations - Page 1 of 16 Groundfish, Offshore Pelagic Species, and Nongame Fish Last saved on 7/27/21 ** NOT ALL REGULATIONS AND DETAILS ARE SHOWN ** Year Effective Jan. 1 (i.e., regulations set preseason) Inseason Change and Effective Date 2021 General marine species: 5-fish 6-fish daily bag limit. 5/10 Daily bag limit is 5 general marine China, copper and quillback rockfishes combined: 1 fish sub- bag limit. species. Cabezon open July 1 – Dec. 31, 1 fish sub-bag limit (of the 5 6 6/1 China, copper and general marine species bag limit), 16” min. quillback rockfishes: Ocean closed seaward of the 40-fathom curve June 1 – Aug. retention prohibited 31. by boats. ________________________________________ Offshore Longleader Fishery: Open outside of the 40-fathom line all year. 10-fish limit. Only these species may be retained: yellowtail, widow, canary, redstripe, greenstriped, silvergray chilipepper, bocaccio, blue and deacon rockfish. Offshore longleader trips cannot be combined with traditional bottomfish, flatfish, or halibut trips except When the bottomfish fishery is restricted to inside of 40- fathoms (June-Aug in 2021) an offshore longleader trip may be combined with all-depth Pacific halibut. However, once rockfish are onboard the vessel, no further halibut fishing may occur. Flatfish Fishery: Open at all depths all year. Daily bag limit: 25 ________________________________________ Retention of yelloweye rockfish is prohibited. Descending devices are mandatory: Any vessel fishing for, or possessing, bottomfish (aka groundfish), including flatfish, or Pacific halibut in the ocean must have a functional descending device onboard, and use it on any rockfish released outside of 30 fathoms. -

US West Coast Rockfish Complex, Cabezon, Kelp Greenling
US West Coast rockfish complex, Cabezon, Kelp Greenling Sebastes spp. and Sebastolobus spp., Scorpaenichthysm armoratus, Hexagrammosdeca grammus Image ©Monterey Bay Aquarium US West Coast Groundfish Fishery (excluding the Pacific hake fishery) Bottom trawl, Midwater trawl, Handline, Bottom longline July 17, 2014 John Driscoll, Consulting researcher 2 About Seafood Watch® The Monterey Bay Aquarium Seafood Watch® program evaluates the ecological sustainability of wild-caught and farmed seafood commonly found in the North American marketplace. Seafood Watch defines sustainable seafood as originating from sources, whether wild-caught or farmed, which can maintain or increase production in the long-term without jeopardizing the structure or function of affected ecosystems. The program’s mission is to engage and empower consumers and businesses to purchase environmentally responsible seafood fished or farmed in ways that minimize their impact on the environment or are in a credible improvement project with the same goal. Each sustainability recommendation is supported by a seafood report. Each report synthesizes and analyzes the most current ecological, fisheries and ecosystem science on a species, then evaluates this information against the program’s sustainability criteria to arrive at a recommendation of “Best Choice,” “Good Alternative,” or “Avoid.” In producing the seafood reports, Seafood Watch utilizes research published in academic, peer-reviewed journals whenever possible. Other sources of information include government technical publications, -

Sebastes Paucispinis) to About 7.4% of Its Former Unfished Population
383 Abstrac t —Although bocaccio (Se- Potential use of offshore marine structures bastes paucispinis) was an economi- cally important rockfish species along in rebuilding an overfished rockfish species, the west coast of North America, overfishing has reduced the stock bocaccio (Sebastes paucispinis) to about 7.4% of its former unfished population. In 2003, using a manned Milton S. Love1 research submersible, we conducted fish surveys around eight oil and gas Donna M. Schroeder1 platforms off southern California as William Lenarz2 part of an assessment of the potential 3 value of these structures as fish habi- Alec MacCall tat. From these surveys, we estimated Ann Scarborough Bull4 that there was a minimum of 430,000 Lyman Thorsteinson5 juvenile bocaccio at these eight struc- tures. We determined this number to 1 Marine Science Institute, University of California be about 20% of the average number Santa Barbara, California, 93160 of juvenile bocaccio that survive annu- E-mail address (for M. S. Love: [email protected] ally for the geographic range of the 2 P.O. 251 species. When these juveniles become Kentfield, California 94914 adults, they will contribute about one percent (0.8%) of the additional 3 NOAA/NMFS Santa Cruz Laboratory amount of fish needed to rebuild the Santa Cruz, California 95060 Pacific Coast population. By compari- 4 U.S. Minerals Management Service son, juvenile bocaccio recruitment to 770 Paseo Camarillo nearshore natural nursery grounds, Camarillo, California 93010 as determined through regional scuba surveys, was low in the same year. 5 Western Fisheries Research Center This research demonstrates that a U.S.