Behavioral Ecology of Crabs: a Breif Review
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

A Classification of Living and Fossil Genera of Decapod Crustaceans
RAFFLES BULLETIN OF ZOOLOGY 2009 Supplement No. 21: 1–109 Date of Publication: 15 Sep.2009 © National University of Singapore A CLASSIFICATION OF LIVING AND FOSSIL GENERA OF DECAPOD CRUSTACEANS Sammy De Grave1, N. Dean Pentcheff 2, Shane T. Ahyong3, Tin-Yam Chan4, Keith A. Crandall5, Peter C. Dworschak6, Darryl L. Felder7, Rodney M. Feldmann8, Charles H. J. M. Fransen9, Laura Y. D. Goulding1, Rafael Lemaitre10, Martyn E. Y. Low11, Joel W. Martin2, Peter K. L. Ng11, Carrie E. Schweitzer12, S. H. Tan11, Dale Tshudy13, Regina Wetzer2 1Oxford University Museum of Natural History, Parks Road, Oxford, OX1 3PW, United Kingdom [email protected] [email protected] 2Natural History Museum of Los Angeles County, 900 Exposition Blvd., Los Angeles, CA 90007 United States of America [email protected] [email protected] [email protected] 3Marine Biodiversity and Biosecurity, NIWA, Private Bag 14901, Kilbirnie Wellington, New Zealand [email protected] 4Institute of Marine Biology, National Taiwan Ocean University, Keelung 20224, Taiwan, Republic of China [email protected] 5Department of Biology and Monte L. Bean Life Science Museum, Brigham Young University, Provo, UT 84602 United States of America [email protected] 6Dritte Zoologische Abteilung, Naturhistorisches Museum, Wien, Austria [email protected] 7Department of Biology, University of Louisiana, Lafayette, LA 70504 United States of America [email protected] 8Department of Geology, Kent State University, Kent, OH 44242 United States of America [email protected] 9Nationaal Natuurhistorisch Museum, P. O. Box 9517, 2300 RA Leiden, The Netherlands [email protected] 10Invertebrate Zoology, Smithsonian Institution, National Museum of Natural History, 10th and Constitution Avenue, Washington, DC 20560 United States of America [email protected] 11Department of Biological Sciences, National University of Singapore, Science Drive 4, Singapore 117543 [email protected] [email protected] [email protected] 12Department of Geology, Kent State University Stark Campus, 6000 Frank Ave. -

National Monitoring Program for Biodiversity and Non-Indigenous Species in Egypt
UNITED NATIONS ENVIRONMENT PROGRAM MEDITERRANEAN ACTION PLAN REGIONAL ACTIVITY CENTRE FOR SPECIALLY PROTECTED AREAS National monitoring program for biodiversity and non-indigenous species in Egypt PROF. MOUSTAFA M. FOUDA April 2017 1 Study required and financed by: Regional Activity Centre for Specially Protected Areas Boulevard du Leader Yasser Arafat BP 337 1080 Tunis Cedex – Tunisie Responsible of the study: Mehdi Aissi, EcApMEDII Programme officer In charge of the study: Prof. Moustafa M. Fouda Mr. Mohamed Said Abdelwarith Mr. Mahmoud Fawzy Kamel Ministry of Environment, Egyptian Environmental Affairs Agency (EEAA) With the participation of: Name, qualification and original institution of all the participants in the study (field mission or participation of national institutions) 2 TABLE OF CONTENTS page Acknowledgements 4 Preamble 5 Chapter 1: Introduction 9 Chapter 2: Institutional and regulatory aspects 40 Chapter 3: Scientific Aspects 49 Chapter 4: Development of monitoring program 59 Chapter 5: Existing Monitoring Program in Egypt 91 1. Monitoring program for habitat mapping 103 2. Marine MAMMALS monitoring program 109 3. Marine Turtles Monitoring Program 115 4. Monitoring Program for Seabirds 118 5. Non-Indigenous Species Monitoring Program 123 Chapter 6: Implementation / Operational Plan 131 Selected References 133 Annexes 143 3 AKNOWLEGEMENTS We would like to thank RAC/ SPA and EU for providing financial and technical assistances to prepare this monitoring programme. The preparation of this programme was the result of several contacts and interviews with many stakeholders from Government, research institutions, NGOs and fishermen. The author would like to express thanks to all for their support. In addition; we would like to acknowledge all participants who attended the workshop and represented the following institutions: 1. -

Belgian Register of Marine Species
BELGIAN REGISTER OF MARINE SPECIES September 2010 Belgian Register of Marine Species – September 2010 BELGIAN REGISTER OF MARINE SPECIES, COMPILED AND VALIDATED BY THE VLIZ BELGIAN MARINE SPECIES CONSORTIUM VLIZ SPECIAL PUBLICATION 46 SUGGESTED CITATION Leen Vandepitte, Wim Decock & Jan Mees (eds) (2010). Belgian Register of Marine Species, compiled and validated by the VLIZ Belgian Marine Species Consortium. VLIZ Special Publication, 46. Vlaams Instituut voor de Zee (VLIZ): Oostende, Belgium. 78 pp. ISBN 978‐90‐812900‐8‐1. CONTACT INFORMATION Flanders Marine Institute – VLIZ InnovOcean site Wandelaarkaai 7 8400 Oostende Belgium Phone: ++32‐(0)59‐34 21 30 Fax: ++32‐(0)59‐34 21 31 E‐mail: [email protected] or [email protected] ‐ 2 ‐ Belgian Register of Marine Species – September 2010 Content Introduction ......................................................................................................................................... ‐ 5 ‐ Used terminology and definitions ....................................................................................................... ‐ 7 ‐ Belgian Register of Marine Species in numbers .................................................................................. ‐ 9 ‐ Belgian Register of Marine Species ................................................................................................... ‐ 12 ‐ BACTERIA ............................................................................................................................................. ‐ 12 ‐ PROTOZOA ........................................................................................................................................... -

DISTRIBUTION PATTERNS of Arenaeus Cribrarius (LAMARCK, 1818) (CRUSTACEA, PORTUNIDAE) in FORTALEZA BA~ UBATUBA (SP), BRAZIL
DISTRIBUTION PATTERNS OF Arenaeus cribrarius (LAMARCK, 1818) (CRUSTACEA, PORTUNIDAE) IN FORTALEZA BA~ UBATUBA (SP), BRAZIL 2 MARCELO ANTONIO AMARO PINHEIRO1,3, ADILSON FRANSOZ0 ,3 2 and MARIA LUCIA NEGREIROS-FRANSOZ0 ,3 lDepartamento de Biologia Aplicada, Faculdade de Ciencias Agnirias e Veterimirias, UNESP, Campus de Iaboticabal - 14870-000 - Iaboticabal, SP, Brasil 2Departamento de Zoologia, Instituto de Biociencias, Cx. Postal 502, UNESP, Cam£us de Botucatu - 18618-000 - Botucatu, SP, Brasil Centro de Aqiiicultura da UNESP (CAUNESP), Nticleo de Estudos em Biologia, Ecologia e Cultivo de Crusmceos (NEBECC) (With 6 figures) ABSTRACT The abundance of the swimming crab A. cribrarius in Fortaleza Bay, Ubatuba (SP) was ana lysed in order to detect the influence of some environmental factors in its distribution. The collections were made by using two otter-trawls deployed from a shrimp-fishing vessel and occurred monthly during one year. The Fortaleza Bay was sampled in 7 radials of I Ian each. Each environmental factor (temperature, salinity and dissolved oxygen of the bottom water, depth, granulometric composition and organic matter of the sediment), sampled in the middle point of each transect, was correlated with the abundance of 5 groups (adult males, ovigerous females, non-ovigerous adult females, juveniles, and total number of specimens), by Pear son's linear and canonical correlation analyses. The total amount of specimens revealed a positive linear correlation with temperature and very fine sand fraction, and a negative linear correlation with organic material contents. Different association patterns appeared in relation to the abundance of the groups mentioned above, such as depth and granulometry. Ovigerous females were the only group which was associated with the whole set of granulometric frac tions of the sediment. -

Evolutionary Transformations of the Reproductive System in Eubrachyura (Crustacea: Decapoda)
EVOLUTIONARY TRANSFORMATIONS OF THE REPRODUCTIVE SYSTEM IN EUBRACHYURA (CRUSTACEA: DECAPODA) DISSERTATION zur Erlangung des akademischen Grades Doctor rerum naturalium (Dr. rer. nat.) eingereicht an der Lebenswissenschaftlichen Fakultät der Humboldt-Universität zu Berlin von M. Sc. Katja, Kienbaum, geb. Jaszkowiak Präsidentin der Humboldt-Universität zu Berlin Prof. Dr.-Ing. Dr. Sabine Kunst Dekan der Lebenswissenschaftlichen Fakultät der Humboldt-Universität zu Berlin Prof. Dr. Bernhard Grimm Gutachter 1. Prof. Dr. Gerhard Scholtz 2. PD Dr. Thomas Stach 3. PD Dr. Christian Wirkner Tag der mündlichen Prüfung: 03.05.2019 CONTENT C ONTENT A BSTRACT v i - vii Z USAMMENFASSUNG viii - x 1 | INTRODUCTION 1 - 11 1.1 | THE BRACHYURA 1 1.1.1 | OBJECT OF INVESTIGATION 1 - 5 1.1.2 | WHAT WE (DO NOT) KNOW ABOUT THE PHYLOGENY OF EUBRACHURA 6 - 10 1. 2 |MS AI 10 - 11 2 | THE MORPHOLOGY OF THE MALE AND FEMALE REPRODUCTIVE SYSTEM IN TWO 12 - 34 SPECIES OF SPIDER CRABS (DECAPODA: BRACHYURA: MAJOIDEA) AND THE ISSUE OF THE VELUM IN MAJOID REPRODUCTION. 2.1 | INTRODUCTION 13 - 14 2.2 | MATERIAL AND METHODS 14 - 16 2.3 | RESULTS 16 - 23 2.4 | DISCUSSION 24 - 34 3 | THE MORPHOLOGY OF THE REPRODUCTIVE SYSTEM IN THE CRAB 35 - 51 PERCNON GIBBESI (DECAPODA: BRACHYURA: GRAPSOIDEA) REVEALS A NEW COMBINATION OF CHARACTERS. 3.1 | INTRODUCTION 36 - 37 3.2 | MATERIAL AND METHODS 37 - 38 3.3 | RESULTS 39 - 46 3.4 | DISCUSSION 46 - 51 4 | THE REPRODUCTIVE SYSTEM OF LIMNOPILOS NAIYANETRI INDICATES A 52 - 64 THORACOTREME AFFILIATION OF HYMENOSOMATIDAE (DECAPODA, EUBRACHYURA). -

Molecular Phylogeny of the Western Atlantic Species of the Genus Portunus (Crustacea, Brachyura, Portunidae)
Blackwell Publishing LtdOxford, UKZOJZoological Journal of the Linnean Society0024-4082The Lin- nean Society of London, 2007? 2007 1501 211220 Original Article PHYLOGENY OF PORTUNUS FROM ATLANTICF. L. MANTELATTO ET AL. Zoological Journal of the Linnean Society, 2007, 150, 211–220. With 3 figures Molecular phylogeny of the western Atlantic species of the genus Portunus (Crustacea, Brachyura, Portunidae) FERNANDO L. MANTELATTO1*, RAFAEL ROBLES2 and DARRYL L. FELDER2 1Laboratory of Bioecology and Crustacean Systematics, Department of Biology, FFCLRP, University of São Paulo (USP), Ave. Bandeirantes, 3900, CEP 14040-901, Ribeirão Preto, SP (Brazil) 2Department of Biology, Laboratory for Crustacean Research, University of Louisiana at Lafayette, Lafayette, LA 70504-2451, USA Received March 2004; accepted for publication November 2006 The genus Portunus encompasses a comparatively large number of species distributed worldwide in temperate to tropical waters. Although much has been reported about the biology of selected species, taxonomic identification of several species is problematic on the basis of strictly adult morphology. Relationships among species of the genus are also poorly understood, and systematic review of the group is long overdue. Prior to the present study, there had been no comprehensive attempt to resolve taxonomic questions or determine evolutionary relationships within this genus on the basis of molecular genetics. Phylogenetic relationships among 14 putative species of Portunus from the Gulf of Mexico and other waters of the western Atlantic were examined using 16S sequences of the rRNA gene. The result- ant molecularly based phylogeny disagrees in several respects with current morphologically based classification of Portunus from this geographical region. Of the 14 species generally recognized, only 12 appear to be valid. -

Invertebrate ID Guide
11/13/13 1 This book is a compilation of identification resources for invertebrates found in stomach samples. By no means is it a complete list of all possible prey types. It is simply what has been found in past ChesMMAP and NEAMAP diet studies. A copy of this document is stored in both the ChesMMAP and NEAMAP lab network drives in a folder called ID Guides, along with other useful identification keys, articles, documents, and photos. If you want to see a larger version of any of the images in this document you can simply open the file and zoom in on the picture, or you can open the original file for the photo by navigating to the appropriate subfolder within the Fisheries Gut Lab folder. Other useful links for identification: Isopods http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-33/htm/doc.html http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-48/htm/doc.html Polychaetes http://web.vims.edu/bio/benthic/polychaete.html http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-34/htm/doc.html Cephalopods http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-44/htm/doc.html Amphipods http://www.19thcenturyscience.org/HMSC/HMSC-Reports/Zool-67/htm/doc.html Molluscs http://www.oceanica.cofc.edu/shellguide/ http://www.jaxshells.org/slife4.htm Bivalves http://www.jaxshells.org/atlanticb.htm Gastropods http://www.jaxshells.org/atlantic.htm Crustaceans http://www.jaxshells.org/slifex26.htm Echinoderms http://www.jaxshells.org/eich26.htm 2 PROTOZOA (FORAMINIFERA) ................................................................................................................................ 4 PORIFERA (SPONGES) ............................................................................................................................................... 4 CNIDARIA (JELLYFISHES, HYDROIDS, SEA ANEMONES) ............................................................................... 4 CTENOPHORA (COMB JELLIES)............................................................................................................................ -

Larval Development of the Speckled Swimming Crab, <I>Arenaeus Cribrarius</I> (Decapoda: Brachyura: Portunidae) Reare
BULLETIN OF MARINE SCIENCE, 42(1): 101-132, 1988 LARVAL DEVELOPMENT OF THE SPECKLED SWIMMING CRAB, ARENAEUS CRIBRARIUS (DECAPODA: BRACHYURA: PORTUNIDAE) REARED IN THE LABORATORY Kenneth C. Stuck and Frank M. Truesdale ABSTRACT Eight zoeal stages and a megalopa of the speckled swimming crab, Arenaeus cribrarius, are described and figured from larvae of an ovigerous specimen collected at Hom Island, Mis- sissippi (Gulf of Mexico). Crab stages 1-3 are also briefly described. Larvae were reared at 25°C and 300/00salinity; some individuals completed zoeal development in 30 days,and reached crab I, 13 days later. Zoea I characters of A. cribrarius are compared with those of seven other species of shallow water Gulf Portuninae, from larvae reared by the authors, and from the literature. Antennal and telson features are important in distinguishing among zoeae I ofGulfPortuninae. Characters ofzoeae II-VIII and the megalopa of A. cribrarius and those of corresponding stages of Callinecles sapidus, C. similis. and PorlUnus spinicarpus from the literature are compared. In zoeae III and VI, setation of maxillar basal and coxal endites is a distinguishing feature among A. cribrarius, C. sapidus, C. similis, and P. spinicarpus, and in zoea V, maxillular basal and coxal endite setation distinguish each. Antennular aesthetasc formula distinguishes each species having a zoea VIII, and in all zoeal stages this feature distinguishes A. cribrarius from the others. Zoeae IV and VII of C. sapidus, C. similis and P. spinicarpus can be separated by setation of maxillular basal and coxal endites, and in zoea II this same feature distinguishes P. spinicarpus from C. -

D Ecapod Crustaceans Associated with the Snakelock Anem One a Nem
SciENTiA M arina71(2) June 2007, 287-292, Barcelona (Spain) ISSN: 0214-8358 Decapod crustaceans associated with the snakelock an em on eAnemonia sulcata. Living there or just passing by? RICARDO CALADO, GISELA DIONÍSIO and MARIA TERESA DINIS CCMAR Universidade do Algarve, Campus de Gambelas, 8000-117 Faro, Portugal. E-mail: [email protected] SUMMARY: The present work identifies the decapod crustaceans that associate with Anemonia sulcata on the southwest ern Atlantic coast of Portugal and characterises their host use pattern. It determines whether the anemone is monopolised by any species, resulting in the exclusion of conspecifics or other decapods and, under laboratory conditions, it evaluates the degree of association between each species and A. sulcata. From all sampled anemones, 79% harboured at least 1 decapod crustacean, with the majority displaying either one or two specimens (32 and 24%, respectively). The most abundant species were the shrimp Periclimenes sagittifer and the crab Inachus phalangium (representing 36 and 31% of collected specimens, respectively), which displayed lasting associations and were commonly recorded among the tentacles of the host. The species Eualus occultus. complexE. cranchii. Clibanarius erythropus. Maja brachydactyla. Pilumnus and hirtellusPolybius (Necora) puber displayed short-term associations, were mainly present on the substratum near the base, and avoided the ten tacles of A. sulcata. Periclimenes sagittifer and I. phalangium were only recorded alone or in heterosexual pairs, appearing to efficiently defend their host against conspecifics. The majority of recorded species only seem to temporarily associate with A. sulcata, in order to seek protection from predators when other shelters are unavailable. Keywords'. Anemonia sulcata, associative behaviour, decapod crustaceans. -

Field Experiments on the Association of Decapod Crustaceans with Sea Anemones, Anemonia Viridis (Forsskål, 1775)
NAT. CROAT. VOL. 19 No 1 151–163 ZAGREB June 30, 2010 original scientific paper/izvorni znanstveni rad FIELD EXPERIMENTS ON THE ASSOCIATION OF DECAPOD CRUSTACEANS WITH SEA ANEMONES, ANEMONIA VIRIDIS (FORSSKÅL, 1775) ROLAND R. MELZER &ROLAND MEYER Zoologische Staatssammlung, Münchhausenstr. 21, D-81247 München, Germany (T: +49898107141, Fax: +49898107300, email: [email protected]) Melzer, R. R. & Meyer, R.: Field experiments on the association of decapod crustaceans with sea anemones, Anemonia viridis (Forsskål, 1775). Nat. Croat., Vol. 19, No. 1, 151–163, 2010, Zagreb. We have undertaken for the first time removal and introduction field experiments with both symbiotic (taken from anemones) and anemone-naïve decapods (taken from somewhere else) and tested defence reactions of putative host anemones. Our findings indicate that decapods go through an individual habituation phase before they become symbionts. Once habituated, they are pro- tected from neighbouring anemones as well. Key words: symbiosis, field experiments, sea anemones, decapods Melzer, R. R. & Meyer, R.: Terenski pokusi na zajednici dekapodnih rakova s vlasuljama, Anemonia viridis (Forsskål, 1775). Nat. Croat., Vol. 19, No. 1, 151–163, 2010, Zagreb. Po prvi puta smo proveli terenske pokuse sa simbiotskim dekapodnim rakovima (uzetim s vlasulja) i onima koji nisu u kontaktu s vlasuljama (uzeti s drugih mjesta) te smo testirali obram- bene reakcije vlasulja-doma}ina. Na{i rezultati pokazuju da dekapodni rakovi prolaze individualnu fazu navikavanja prije nego {to postanu simbionti. Nakon toga budu za{ti}eni i od susjednih vlasulja. Klju~ne rije~i: simbioza, terenski pokusi, moruzgve i vlasulje, dekapodni rakovi INTRODUCTION Decapod crustaceans living as symbionts, commensals or ectoparasites on sea anemones and other cnidarian hosts is a common phenomenon observed for nu- merous species of decapods as well as cnidarians (WEINBAUER et al., 1982; WIRTZ & DIESEL, 1983; ROSS, 1983; FAUTIN et al., 1995). -
St. Lucie, Units 1 and 2
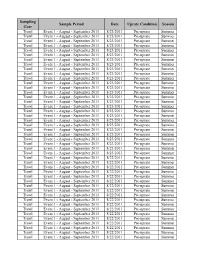
Sampling Sample Period Date Uprate Condition Season Gear Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event 1 - August - September 2011 8/23/2011 Pre-uprate Summer Trawl Event -

Feeding and Masking Selection in Inachus Phalangium (Decapoda, Majidae): Dressing up Has Never Been So Complicated
MARINE ECOLOGY PROGRESS SERIES Vol. 336: 225–233, 2007 Published April 27 Mar Ecol Prog Ser Feeding and masking selection in Inachus phalangium (Decapoda, Majidae): dressing up has never been so complicated Rocco Rorandelli*, Marina Gomei, Marco Vannini, Stefano Cannicci Dipartimento di Biologia Animale e Genetica ‘Leo Pardi’, Università degli Studi di Firenze, Via Romana 17, 50125 Firenze, Italy ABSTRACT: We investigated the choice preferences exhibited by the spider crab Inachus pha- langium when using algae for feeding and masking, together with the pattern of decoration present on the body surface and the morphology of the setae, which are used to attach masking material. We showed that feeding and masking preferences are decoupled, and that I. phalangium exploited the algal nutritional and chemical properties differently. When feeding, the crab revealed a differential choice among the offered algae, preferring the protein-rich cyanobacteria. When decorating, both algal components that match the background and the Phaeophyta Dictyota dichotoma, which acts as a chemical deterrent against predators, were actively chosen. At the same time, decoration in I. pha- langium was used as a short-term food store. We also provided evidence that masking patterns varied in different body regions, depending on the functional morphology of the hooked setae. This work discusses the origin and evolution of the decorative habit, showing for the first time that decoration is a multifaceted mechanism, tightly linked with morpho-functional and behavioural adaptations, and that masking material can also be used as a food store. KEY WORDS: Chemically mediated defence · Decoration · Dictyota · Functional morphology · Spider crab Resale or republication not permitted without written consent of the publisher INTRODUCTION (Wicksten 1980, 1983, 1993, Woods & McLay 1994).