Embryonic Growth and the Evolution of the Selfish V-Linked Growth Factors
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

ZFX Acts As a Transcriptional Activator in Multiple Types of Human Tumors by Binding Downstream from Transcription Start Sites at the Majority of Cpg Island Promoters
Downloaded from genome.cshlp.org on October 6, 2021 - Published by Cold Spring Harbor Laboratory Press Research ZFX acts as a transcriptional activator in multiple types of human tumors by binding downstream from transcription start sites at the majority of CpG island promoters Suhn Kyong Rhie, Lijun Yao, Zhifei Luo, Heather Witt, Shannon Schreiner, Yu Guo, Andrew A. Perez, and Peggy J. Farnham Department of Biochemistry and Molecular Medicine and the Norris Comprehensive Cancer Center, Keck School of Medicine, University of Southern California, Los Angeles, California 90089, USA High expression of the transcription factor ZFX is correlated with proliferation, tumorigenesis, and patient survival in mul- tiple types of human cancers. However, the mechanism by which ZFX influences transcriptional regulation has not been determined. We performed ChIP-seq in four cancer cell lines (representing kidney, colon, prostate, and breast cancers) to identify ZFX binding sites throughout the human genome. We identified roughly 9000 ZFX binding sites and found that most of the sites are in CpG island promoters. Moreover, genes with promoters bound by ZFX are expressed at higher levels than genes with promoters not bound by ZFX. To determine if ZFX contributes to regulation of the promoters to which it is bound, we performed RNA-seq analysis after knockdown of ZFX by siRNA in prostate and breast cancer cells. Many genes with promoters bound by ZFX were down-regulated upon ZFX knockdown, supporting the hypothesis that ZFX acts as a transcriptional activator. Surprisingly, ZFX binds at +240 bp downstream from the TSS of the responsive pro- moters. Using Nucleosome Occupancy and Methylome Sequencing (NOMe-seq), we show that ZFX binds between the open chromatin region at the TSS and the first downstream nucleosome, suggesting that ZFX may play a critical role in pro- moter architecture. -

Sequence Analysis in Bos Taurus Reveals Pervasiveness of X–Y Arms Races in Mammalian Lineages
Downloaded from genome.cshlp.org on September 25, 2021 - Published by Cold Spring Harbor Laboratory Press Research Sequence analysis in Bos taurus reveals pervasiveness of X–Y arms races in mammalian lineages Jennifer F. Hughes,1 Helen Skaletsky,1,2 Tatyana Pyntikova,1 Natalia Koutseva,1 Terje Raudsepp,3 Laura G. Brown,1,2 Daniel W. Bellott,1 Ting-Jan Cho,1 Shannon Dugan-Rocha,4 Ziad Khan,4 Colin Kremitzki,5 Catrina Fronick,5 Tina A. Graves-Lindsay,5 Lucinda Fulton,5 Wesley C. Warren,5,7 Richard K. Wilson,5,8 Elaine Owens,3 James E. Womack,3 William J. Murphy,3 Donna M. Muzny,4 Kim C. Worley,4 Bhanu P. Chowdhary,3,9 Richard A. Gibbs,4 and David C. Page1,2,6 1Whitehead Institute, Cambridge, Massachusetts 02142, USA; 2Howard Hughes Medical Institute, Whitehead Institute, Cambridge, Massachusetts 02142, USA; 3College of Veterinary Medicine and Biomedical Sciences, Texas A&M University, College Station, Texas 77843, USA; 4Human Genome Sequencing Center, Baylor College of Medicine, Houston, Texas 77030, USA; 5The McDonnell Genome Institute, Washington University School of Medicine, St. Louis, Missouri 63108, USA; 6Department of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02142, USA Studies of Y Chromosome evolution have focused primarily on gene decay, a consequence of suppression of crossing-over with the X Chromosome. Here, we provide evidence that suppression of X–Y crossing-over unleashed a second dynamic: selfish X–Y arms races that reshaped the sex chromosomes in mammals as different as cattle, mice, and men. Using su- per-resolution sequencing, we explore the Y Chromosome of Bos taurus (bull) and find it to be dominated by massive, lin- eage-specific amplification of testis-expressed gene families, making it the most gene-dense Y Chromosome sequenced to date. -

Differential Expression of ZFX Gene in Gastric Cancer
Differential expression of ZFX gene in gastric cancer 1 2,3, 1 PARVANEH NIKPOUR , MODJTABA EMADI-BAYGI *, FAEZEH MOHAMMAD-HASHEM , MOHAMAD REZA MARACY4 and SHAGHAYEGH HAGHJOOY-JAVANMARD5 1Department of Genetics and Molecular Biology, Faculty of Medicine, Isfahan University of Medical Sciences, Isfahan, Iran 2Department of Genetics, Faculty of Basic Sciences, 3Research Institute of Biotechnology, Shahrekord University, Shahrekord, Iran 4Department of Community Medicine, Faculty of Medicine, 5Applied Physiology Research Center, Isfahan University of Medical Sciences, Isfahan, Iran *Corresponding author (Fax, +98-311-7753480; Email, [email protected]) Gastric cancer accounts for 8% of the total cancer cases and 10% of total cancer deaths worldwide. In Iran, gastric cancer is the leading cause of national cancer-related mortality. Most human cancers show substantial heterogeneity. The cancer stem cell (CSC) hypothesis has been proposed to reconcile this heterogeneity. ZFX encodes a member of the krueppel C2H2-type zinc-finger protein family that is required as a transcriptional regulator for self-renewal of stem cells. A total of 30 paired tissue gastric samples were examined for ZFX gene expression by quantitative real-time RT-PCR. Although the relative expression of the gene was significantly high in 47% of the examined tumour tissues, its expression was low in the others (53%). There was a statistically significant association between the ZFX gene expression and different tumour types and grades. This is the first report that shows ZFX was differentially expressed in gastric cancer. Of note, it was overexpressed in diffused-type and grade III gastric tumoural tissues. Due to this, ZFX may have the potential to be used as a target for therapeutic interventions. -

Genetics of Azoospermia
International Journal of Molecular Sciences Review Genetics of Azoospermia Francesca Cioppi , Viktoria Rosta and Csilla Krausz * Department of Biochemical, Experimental and Clinical Sciences “Mario Serio”, University of Florence, 50139 Florence, Italy; francesca.cioppi@unifi.it (F.C.); viktoria.rosta@unifi.it (V.R.) * Correspondence: csilla.krausz@unifi.it Abstract: Azoospermia affects 1% of men, and it can be due to: (i) hypothalamic-pituitary dysfunction, (ii) primary quantitative spermatogenic disturbances, (iii) urogenital duct obstruction. Known genetic factors contribute to all these categories, and genetic testing is part of the routine diagnostic workup of azoospermic men. The diagnostic yield of genetic tests in azoospermia is different in the different etiological categories, with the highest in Congenital Bilateral Absence of Vas Deferens (90%) and the lowest in Non-Obstructive Azoospermia (NOA) due to primary testicular failure (~30%). Whole- Exome Sequencing allowed the discovery of an increasing number of monogenic defects of NOA with a current list of 38 candidate genes. These genes are of potential clinical relevance for future gene panel-based screening. We classified these genes according to the associated-testicular histology underlying the NOA phenotype. The validation and the discovery of novel NOA genes will radically improve patient management. Interestingly, approximately 37% of candidate genes are shared in human male and female gonadal failure, implying that genetic counselling should be extended also to female family members of NOA patients. Keywords: azoospermia; infertility; genetics; exome; NGS; NOA; Klinefelter syndrome; Y chromosome microdeletions; CBAVD; congenital hypogonadotropic hypogonadism Citation: Cioppi, F.; Rosta, V.; Krausz, C. Genetics of Azoospermia. 1. Introduction Int. J. Mol. Sci. -

ZFX Rabbit Polyclonal Antibody – TA343385 | Origene
OriGene Technologies, Inc. 9620 Medical Center Drive, Ste 200 Rockville, MD 20850, US Phone: +1-888-267-4436 [email protected] EU: [email protected] CN: [email protected] Product datasheet for TA343385 ZFX Rabbit Polyclonal Antibody Product data: Product Type: Primary Antibodies Applications: WB Recommended Dilution: WB Reactivity: Human Host: Rabbit Isotype: IgG Clonality: Polyclonal Immunogen: The immunogen for Anti-ZFX antibody is: synthetic peptide directed towards the C-terminal region of Human ZFX. Synthetic peptide located within the following region: TTDASGFKRHVISIHTKDYPHRCEYCKKGFRRPSEKNQHIMRHHKEVGLP Formulation: Liquid. Purified antibody supplied in 1x PBS buffer with 0.09% (w/v) sodium azide and 2% sucrose. Note that this product is shipped as lyophilized powder to China customers. Purification: Affinity Purified Conjugation: Unconjugated Storage: Store at -20°C as received. Stability: Stable for 12 months from date of receipt. Predicted Protein Size: 83 kDa Gene Name: zinc finger protein, X-linked Database Link: NP_003401 Entrez Gene 7543 Human P17010 Background: The function of this protein remains unknown. Synonyms: ZNF926 Note: Immunogen Sequence Homology: Dog: 100%; Pig: 100%; Rat: 100%; Horse: 100%; Human: 100%; Mouse: 100%; Bovine: 100%; Rabbit: 100%; Zebrafish: 100%; Guinea pig: 100% Protein Families: Transcription Factors This product is to be used for laboratory only. Not for diagnostic or therapeutic use. View online » ©2021 OriGene Technologies, Inc., 9620 Medical Center Drive, Ste 200, Rockville, MD 20850, US 1 / 2 ZFX Rabbit Polyclonal Antibody – TA343385 Product images: Host: Rabbit; Target Name: ZFX; Sample Tissue: ACHN Whole cell lysates; Antibody Dilution: 1.0 ug/ml This product is to be used for laboratory only. -

Molecular Evolution of a Y Chromosome to Autosome Gene Duplication in Drosophila Research Article
Molecular Evolution of a Y Chromosome to Autosome Gene Duplication in Drosophila Kelly A. Dyer,*,1 Brooke E. White,1 Michael J. Bray,1 Daniel G. Pique´,1 and Andrea J. Betancourt* ,2 1Department of Genetics, University of Georgia 2Institute of Evolutionary Biology, University of Edinburgh, Ashworth Labs, Edinburgh, United Kingdom Present address: Institute for Population Genetics, University of Veterinary Medicine Vienna, Vienna 1210, Austria *Corresponding author: [email protected], [email protected]. Associate editor: Jody Hey Abstract In contrast to the rest of the genome, the Y chromosome is restricted to males and lacks recombination. As a result, Research article Y chromosomes are unable to respond efficiently to selection, and newly formed Y chromosomes degenerate until few genes remain. The rapid loss of genes from newly formed Y chromosomes has been well studied, but gene loss from highly degenerate Y chromosomes has only recently received attention. Here, we identify and characterize a Y to autosome duplication of the male fertility gene kl-5 that occurred during the evolution of the testacea group species of Drosophila. The duplication was likely DNA based, as other Y-linked genes remain on the Y chromosome, the locations of introns are conserved, and expression analyses suggest that regulatory elements remain linked. Genetic mapping reveals that the autosomal copy of kl-5 resides on the dot chromosome, a tiny autosome with strongly suppressed recombination. Molecular evolutionary analyses show that autosomal copies of kl-5 have reduced polymorphism and little recombination. Importantly, the rate of protein evolution of kl-5 has increased significantly in lineages where it is on the dot versus Y linked. -

©Copyright 2008 Joseph A. Ross the Evolution of Sex-Chromosome Systems in Stickleback Fishes
©Copyright 2008 Joseph A. Ross The Evolution of Sex-Chromosome Systems in Stickleback Fishes Joseph A. Ross A dissertation submitted in partial fulfillment of the requirements for the degree of Doctor of Philosophy University of Washington 2008 Program Authorized to Offer Degree: Molecular and Cellular Biology University of Washington Graduate School This is to certify that I have examined this copy of a doctoral dissertation by Joseph A. Ross and have found that it is complete and satisfactory in all respects, and that any and all revisions required by the final examining committee have been made. Chair of the Supervisory Committee: Catherine L. Peichel Reading Committee: Catherine L. Peichel Steven Henikoff Barbara J. Trask Date: In presenting this dissertation in partial fulfillment of the requirements for the doctoral degree at the University of Washington, I agree that the Library shall make its copies freely available for inspection. I further agree that extensive copying of the dissertation is allowable only for scholarly purposes, consistent with “fair use” as prescribed in the U.S. Copyright Law. Requests for copying or reproduction of this dissertation may be referred to ProQuest Information and Learning, 300 North Zeeb Road, Ann Arbor, MI 48106-1346, 1-800-521-0600, to whom the author has granted “the right to reproduce and sell (a) copies of the manuscript in microform and/or (b) printed copies of the manuscript made from microform.” Signature Date University of Washington Abstract The Evolution of Sex-Chromosome Systems in Stickleback Fishes Joseph A. Ross Chair of the Supervisory Committee: Affiliate Assistant Professor Catherine L. -

Evolution of Y Chromosome Ampliconic Genes in Great Apes
The Pennsylvania State University The Graduate School EVOLUTION OF Y CHROMOSOME AMPLICONIC GENES IN GREAT APES A Dissertation in Bioinformatics and Genomics by Rahulsimham Vegesna © 2020 Rahulsimham Vegesna Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy May 2020 The dissertation of Rahulsimham Vegesna was reviewed and approved by the following: Paul Medvedev Associate Professor of Computer Science & Engineering Associate Professor of Biochemistry & Molecular Biology Dissertation Co-Adviser Co-Chair of Committee Kateryna D. Makova Pentz Professor of Biology Dissertation Co-Adviser Co-Chair of Committee Michael DeGiorgio Associate Professor of Biology and Statistics Wansheng Liu Professor of Animal Genomics George H. Perry Chair, Intercollege Graduate Degree Program in Bioinformatics and Genomics Associate Professor of Anthropology and Biology ii ABSTRACT In addition to the sex-determining gene SRY and several other single-copy genes, the human Y chromosome harbors nine multi-copy gene families which are expressed exclusively in testis. In humans, these gene families are important for spermatogenesis and their loss is observed in patients suffering from infertility. However, only five of the nine ampliconic gene families are found across great apes, while others are missing or pseudogenized in some species. My research goal is to understand the evolution of the Y ampliconic gene families in humans and in non-human great ape species. The specific objectives I addressed in this dissertation are 1. To test whether Y ampliconic gene expression levels depend on their copy number and whether there is a gene dosage compensation to counteract the ampliconic gene copy number variation observed in humans. -
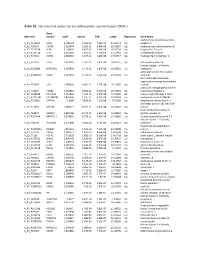
Table S1. the Statistical Metrics for Key Differentially Expressed Genes (Degs)
Table S1. The statistical metrics for key differentially expressed genes (DEGs) Gene Agilent Id Symbol logFC pValue FDR tvalue Regulation Gene Name oxidized low density lipoprotein A_24_P124624 OLR1 2.458429 1.19E-13 7.25E-10 24.04241 Up receptor 1 A_23_P90273 CHST8 2.622464 3.85E-12 6.96E-09 19.05867 Up carbohydrate sulfotransferase 8 A_23_P217528 KLF8 2.109007 4.85E-12 7.64E-09 18.76234 Up Kruppel like factor 8 A_23_P114740 CFH 2.651636 1.85E-11 1.79E-08 17.13652 Up complement factor H A_23_P34031 XAGE2 2.000935 2.04E-11 1.81E-08 17.02457 Up X antigen family member 2 A_23_P27332 TCF4 1.613097 2.32E-11 1.87E-08 16.87275 Up transcription factor 4 histone cluster 1 H1 family A_23_P250385 HIST1H1B 2.298658 2.47E-11 1.87E-08 16.80362 Up member b abnormal spindle microtubule A_33_P3288159 ASPM 2.162032 2.79E-11 2.01E-08 16.66292 Up assembly H19, imprinted maternally expressed transcript (non-protein A_24_P52697 H19 1.499364 4.09E-11 2.76E-08 16.23387 Up coding) potassium voltage-gated channel A_24_P31627 KCNB1 2.289689 6.65E-11 3.97E-08 15.70253 Up subfamily B member 1 A_23_P214168 COL12A1 2.155835 7.59E-11 4.15E-08 15.56005 Up collagen type XII alpha 1 chain A_33_P3271341 LOC388282 2.859496 7.61E-11 4.15E-08 15.55704 Up uncharacterized LOC388282 A_32_P150891 DIAPH3 2.2068 7.83E-11 4.22E-08 15.5268 Up diaphanous related formin 3 zinc finger protein 185 with LIM A_23_P11025 ZNF185 1.385721 8.74E-11 4.59E-08 15.41041 Up domain heat shock protein family B A_23_P96872 HSPB11 1.887166 8.94E-11 4.64E-08 15.38599 Up (small) member 11 A_23_P107454 -

Genetics, a Mating Experiment Called a Reciprocal Cross Is Performed to Test If an Animal's Trait Is Sex-Linked
Sex Determination Sex determination, is a system which decides the sexual characteristics of an organism or offspring. It helps to determine whether the organism will be male or a female, which are the two most common sexes. The widely used technique is chromosomal sex determination, in which sex chromosome of male i.e. X or Y chromosome, decides the sex or gender of the offspring. Female carries XX chromosome and male carries XY chromosome. It is also possible to perform genetic tests to eliminate any chromosomal or genetic disorders. Sex determination is the genetic process of determining the sex of the organism. Let us study in more detail about this. Before we understand how sex determination is done, we need to understand the genetic makeup of a human being. As we all know, humans have 23 pairs or 46 chromosomes. Of these 23 pairs, 22 pairs are known as autosomes whereas 1 pair is known as the sex chromosome. It is this one pair that helps in determining the sex of an individual. Sex Determination in Humans Females in humans have 2X chromosomes- 1 each is inherited from either parent and denoted as XX. Males in humans have 1X and 1 Y chromosome, where the X is inherited from the mother and the Y from the father. In a way, we can say that it is the father who determines the sex of the unborn child. This can put to shame a lot of history in which people believed it is the woman who was responsible for not giving birth to a male heir. -

Cpg Island-Mediated Global Gene Regulatory Modes in Mouse Embryonic Stem Cells
ARTICLE Received 26 Mar 2014 | Accepted 3 Oct 2014 | Published 18 Nov 2014 DOI: 10.1038/ncomms6490 OPEN CpG island-mediated global gene regulatory modes in mouse embryonic stem cells Samuel Beck1, Bum-Kyu Lee1, Catherine Rhee1, Jawon Song2, Andrew J. Woo3 & Jonghwan Kim1,4,5 Both transcriptional and epigenetic regulations are fundamental for the control of eukaryotic gene expression. Here we perform a compendium analysis of 4200 large sequencing data sets to elucidate the regulatory logic of global gene expression programs in mouse embryonic stem (ES) cells. We define four major classes of DNA-binding proteins (Core, PRC, MYC and CTCF) based on their target co-occupancy, and discover reciprocal regulation between the MYC and PRC classes for the activity of nearly all genes under the control of the CpG island (CGI)-containing promoters. This CGI-dependent regulatory mode explains the functional segregation between CGI-containing and CGI-less genes during early development. By defining active enhancers based on the co-occupancy of the Core class, we further demon- strate their additive roles in CGI-containing gene expression and cell type-specific roles in CGI-less gene expression. Altogether, our analyses provide novel insights into previously unknown CGI-dependent global gene regulatory modes. 1 Department of Molecular Biosciences, The University of Texas at Austin, Austin, Texas 78712, USA. 2 Texas Advanced Computing Center, The University of Texas at Austin, Austin, Texas 78758, USA. 3 School of Medicine and Pharmacology, Royal Perth Hospital Unit, The University of Western Australia, Perth, WA 6000, Australia. 4 Institute for Cellular and Molecular Biology, The University of Texas at Austin, Austin, Texas 78712, USA. -

Expression of ZFX Gene Correlated with the Central Features of the Neoplastic Phenotype in Human Brain Tumors with Distinct Phenotypes
Original Article Expression of ZFX gene correlated with the central features of the neoplastic phenotype in human brain tumors with distinct phenotypes Azita Afzali, Modjtaba Emadi‑Baygi, Parvaneh Nikpour1, Fatemehe Nazemroaya1, Majid Kheirollahi1 Department of Genetics, Faculty of Basic Sciences, Shahrekord University, Shahrekord, 1Department of Genetics and Molecular Biology, School of Medicine, Pediatric Inherited Diseases Research Center, Research Institute for Primordial Prevention of Non-communicable Disease, Isfahan University of Medical Sciences, Isfahan, Iran Abstract Background: The zinc finger transcription factor zinc finger protein, X‑linked (ZFX) acts as an important director of self‑renewal in several stem cell types. Moreover, ZFX expression abnormally increases in various cancers and relates to tumor grade. We performed this study, to examine its role in the pathogenesis of astrocytoma and meningioma. Materials and Methods: We used real‑time reverse transcription polymerase chain reaction method for evaluation of ZFX expression in 25 astrocytoma tumoral tissue and 25 meningioma tumoral tissues with different WHO grades. Furthermore, the association of gene expression with various clinic‑pathological characteristics was examined. Results: We found that there is a significant association between gene expression and different tumor grades, the presence or absence of invasion, forming and nonforming of glomeruloid vessels, the age over or under 50 and the presence or absence of calcification in astrocytomas. This is the first report that shows that ZFX was directly correlated with the central features of the neoplastic phenotype, including the growth of cancer cells, angiogenesis, and invasion. Conclusion: Regarding all the above‑mentioned studies, it is highly plausible that silencing the expression of ZFX gene in gliomas has a major role in the therapeutic interventions of the disease in future.